
Plan de gestion de l’otarie de Steller (Eumetopias jubatus) au Canada 2010

La série des plans de gestion de la Loi sur les espèces en péril
Qu’est-ce que la Loi sur les espèces en péril (LEP)?
La LEP est la loi fédérale qui constitue l’une des pierres d’assise de l’effort national commun de protection et de conservation des espèces en péril au Canada. La Loi est en vigueur depuis 2003 et vise, entre autres, à « favoriser la gestion des espèces préoccupantes pour éviter qu’elles ne deviennent des espèces en voie de disparition ou menacées ».
Qu’est-ce qu’une espèce préoccupante?
Selon la LEP, une espèce préoccupante est une espèce sauvage qui peut devenir une espèce menacée ou une espèce en voie de disparition par l’effet cumulatif de ses caractéristiques biologiques et des menaces signalées à son égard. Les espèces préoccupantes sont inscrites à la Liste des espèces en péril de la LEP.
Qu’est-ce qu’un plan de gestion?
Selon la LEP, un plan de gestion est un document de planification axé sur l’action qui désigne les activités de conservation et les mesures relatives à l’utilisation des terres qu’il faut prendre pour éviter, à tout le moins, que l’espèce préoccupante ne devienne menacée ou en voie de disparition. Pour de nombreuses espèces, le but ultime d’un plan de gestion est d’atténuer les menaces d’origine anthropique et de retirer l’espèce de la Liste des espèces en péril. Le plan fixe des buts et des objectifs, identifie les menaces et propose les principales activités à entreprendre pour atténuer ces dernières.
L’élaboration de plans de gestion est obligatoire en vertu des articles 65 à 72 de la LEP
(http://www.registrelep.gc.ca/default_f.cfm).
Le plan de gestion doit être préparé au plus tard trois ans après l’inscription de l’espèce à la Liste des espèces en péril. Dans le cas des espèces qui ont été inscrites à la LEP lorsque celle-ci a été adoptée, le délai est de cinq ans.
Et ensuite?
Les orientations contenues dans le plan de gestion permettront aux entités responsables, aux collectivités, aux utilisateurs des terres et aux conservationnistes de mettre en œuvre des mesures de conservation qui auront des effets préventifs ou réparateurs. Le manque de certitude scientifique ne doit pas servir de prétexte pour retarder la prise de mesures efficaces pour éviter qu’une espèce ne devienne davantage en péril; la mise en œuvre de telles mesures pourrait même éviter d’importantes dépenses dans le futur.
La série des plans de gestion
Cette série présente les plans de gestion élaborés ou adoptés par le gouvernement fédéral dans le cadre de la LEP. De nouveaux documents s’ajouteront régulièrement à mesure que de nouvelles espèces seront inscrites et que les plans de gestion actuels seront mis à jour.
Pour en savoir davantage
Pour en savoir davantage sur la Loi sur les espèces en péril et les initiatives de conservation, veuillez consulter le Registre public de la LEP (http://www.registrelep.gc.ca).
Référence recommandée :
Pêches et Océans Canada. 2010. Plan de gestion de l’otarie de Steller (Eumetopias jubatus ) au Canada [Proposition]. Série des plans de gestion de la Loi sur les espèces en péril. Pêches et Océans Canada, Ottawa. vii + 80 p.
Exemplaires supplémentaires :
Il est possible de télécharger des exemplaires de la présente publication à partir du Registre public des espèces en péril : (http://www.registrelep.gc.ca/).
Illustration de la couverture : M.A. Bigg.
Also available in English under the title:
“Proposed Management Plan for the Steller Sea Lion (Eumetopias jubatus) in Canada”
© Sa Majesté la Reine du Chef du Canada, représentée par le ministre des Pêches et des Océans du Canada, 2010. Tous droits réservés.
ISBN 978-1-100-94977-2
No de catalogue : En3-5/8-2010F-PDF
Le contenu du présent document (sauf les illustrations) peut être utilisé sans permission, à condition que la source soit adéquatement citée.
L’otarie de Steller est un mammifère marin qui relève de la compétence du gouvernement fédéral. Le ministre des Pêches et des Océans est un « ministre compétent » en matière d’espèces aquatiques en vertu de la Loi sur les espèces en péril (LEP). Puisque les otaries de Steller vivent dans la réserve de parc national et le site du patrimoine haïda Gwaii Haanas, ainsi que dans la réserve de parc national Pacific Rim et dans la réserve de parc national des Îles-Gulf, qui sont administrées par l’Agence Parcs Canada, le ministre de l’Environnement est également un « ministre compétent » en vertu de la LEP pour les individus de l’espèce présents sur les terres et dans les eaux dont la gestion relève de Parcs Canada. La LEP (articles 65 et 66) stipule que les ministres compétents doivent préparer des plans de gestion pour les espèces inscrites comme étant préoccupantes, en collaboration et en consultation avec les parties intéressées et touchées. L’otarie de Steller a été inscrite en tant qu’espèce préoccupante en vertu de la LEP en juillet 2005. Pêches et Océans Canada, Région du Pacifique, a dirigé l’élaboration du présent plan de gestion, en collaboration et en consultation avec un grand nombre de personnes, d’organisations et d’organismes gouvernementaux, y compris l’Agence Parcs Canada (annexe III).
La réussite de la conservation de cette espèce dépendra de l’engagement et de la collaboration d’un grand nombre de parties qui participeront à la mise en œuvre des orientations formulées dans le présent plan et ne pourra reposer sur Pêches et Océans Canada, sur l’Agence Parcs Canada ou sur une autre instance seulement. Le présent plan vise à guider les administrations et les organismes qui participent ou qui souhaitent participer aux activités de conservation de l’espèce. Dans l’esprit de l’Accord national pour la protection des espèces en péril, le ministre des Pêches et des Océans et le ministre de l’Environnement invitent toutes les entités responsables ainsi que tous les Canadiens à se joindre à Pêches et Océans Canada et à l’Agence Parcs Canada pour appuyer le présent plan et le mettre en œuvre au profit de l’otarie de Steller et de l’ensemble de la société canadienne. Les ministres compétents rendront compte des progrès réalisés d’ici cinq ans.
Pêches et Océans Canada
Gouvernement de la Colombie-Britannique
Environnement Canada
Agence Parcs Canada
L’équipe technique du MPO a rédigé le présent document pour Pêches et Océans Canada (MPO).
L’équipe aimerait remercier Alana Phillips pour sa contribution à l’élaboration des premières versions ainsi que pour la coordination et l’organisation de l’atelier technique. Pêches et Océans Canada désire également souligner la participation de toutes les personnes qui ont assisté à l’atelier technique sur la planification de la gestion de la population d’otaries de Steller (voir l’annexe III). Le compte rendu de l’atelier a permis la formulation d’un avis technique et scientifique utile concernant les déplacements des otaries de Steller, leur répartition et leur abondance, lequel avis a contribué à la préparation du présent document.
Conformément à la Directive du Cabinet sur l’évaluation environnementale des projets de politiques, de plans et de programmes, une évaluation environnementale stratégique (EES) doit être menée pour tous les documents de planification du rétablissement produits en vertu de la LEP. L’objet de l’EES est d’intégrer les considérations environnementales à l’élaboration des projets de politiques, de plans et de programmes publics afin de soutenir la prise de décisions éclairées sur le plan environnemental.
La planification de la gestion profitera aux espèces en péril et à la biodiversité en général. Il est toutefois reconnu que des plans peuvent produire, sans que cela ne soit voulu, des effets environnementaux négatifs qui dépassent les avantages prévus. Le processus de planification fondé sur des lignes directrices nationales tient directement compte de tous les effets environnementaux, notamment des impacts possibles sur les espèces ou les habitats non ciblés. Les résultats de l’EES sont directement intégrés au plan de gestion même, mais sont également résumés ci-après.
Tout au long du processus d’élaboration du présent plan, on a procédé à l’évaluation de nombreux facteurs qui mettent en péril ou qui risquent de mettre en péril la gestion de la population d’otaries de Steller, lesquels facteurs sont présentés ci-après. Parmi les principaux facteurs ou menaces anthropiques, mentionnons la concurrence exercée par les pêches et la contamination par les produits chimiques. La variabilité environnementale qui limite la disponibilité des proies est également un facteur préoccupant concernant les impacts à long terme sur la santé de la population. Dans certains cas, ces facteurs menacent les populations, tandis que dans d’autres cas, ils affectent l’habitat désigné. Il ressort de l’exercice que certaines menaces peuvent être atténuées par l’application des lois, des politiques et des programmes en vigueur et, en fait, on recense de nombreux exemples de mesures d’atténuation qui sont mises en œuvre actuellement. Cependant, dans d’autres cas, la menace ou les mesures d’atténuation potentielles doivent faire l’objet d’autres travaux de recherche ou d’une évaluation avant que l’on puisse formuler des recommandations sur des mesures ou des activités particulières. Le type général des travaux de recherche, d’évaluations et d’approches qui sont nécessaires à l’atténuation de ces menaces est décrit dans le présent plan de gestion.
Pendant le processus de mise en œuvre des mesures, des activités précises en matière de gestion, de rétablissement et d’atténuation des menaces seront évaluées et exposées en détail pour ces populations et seront accompagnées d’une évaluation des effets et des coûts de chaque activité ou mesure. En conséquence, si nous tenons compte de la nature générale des nouvelles mesures d’atténuation recommandées pour la gestion de ces populations et du fait que nombre des recommandations visant à protéger l’habitat relèvent de lois et de politiques en vigueur, nous pouvons affirmer que le présent plan n’entraînera pas de nouveaux effets négatifs importants.
L’otarie de Steller, le plus grand représentant des phoques à oreilles, est une espèce relativement longévive à reproduction lente. Son nom scientifique, Eumetopias jubatus, signifie « qui a un large front et une crinière » et fait référence au collier proéminent de poils drus que portent les mâles adultes autour du cou et de la poitrine et qui ressemble à la crinière d’un lion.
Toutes les otaries de Steller sont remarquablement agiles sur terre en raison de leur capacité de tourner leurs nageoires postérieures vers l’avant et de s’appuyer sur leurs nageoires antérieures. Les otaries de Steller peuvent grimper sur des rochers escarpés et sont souvent observées à de nombreux mètres au-dessus de la surface de la mer. Sur terre, elles ont tendance à être très grégaires et se rassemblent en groupes compacts dans des colonies de reproduction denses (roqueries) ou sur des échoueries ne servant pas à la reproduction. Bien que les otaries de Steller fréquentent habituellement des échoueries de façon régulière, elles passent parfois de nombreux jours ou plusieurs semaines en mer sans venir à terre et peuvent dormir dans l’eau, habituellement dans des groupes que l’on appelle des « radeaux ».
La répartition de cette espèce est transfrontalière, et les activités de recherche et de gestion réalisées à son égard sont actuellement menées par les gouvernements canadien et américain de même que par des organismes privés et des chercheurs indépendants. On recense actuellement quatre sites de reproduction en Colombie-Britannique, dont l’un a été repeuplé récemment après avoir été déserté à la suite des programmes d’abattage des prédateurs dans les années 1920. L’échouerie des îles Scott, en particulier, figure au deuxième rang en importance au monde parmi les concentrations de reproduction, et la Colombie-Britannique soutient actuellement environ 33 % de l’effectif total de la population d’otaries de Steller de l’Est. En C.-B., les otaries de Steller se reproduisent actuellement à toutes les roqueries historiques connues, et les populations ont maintenant largement dépassé les sommets historiques connus.
Selon l’estimation pour la production de petits dérivée du dernier relevé mené sur l’ensemble de l’aire de répartition en 2002, la taille totale de la population de l’Est oscillerait entre 46 000 et 58 000 individus. On a estimé que l’abondance dans les eaux canadiennes était de 20 000 à 28 000 individus selon le relevé le plus récent mené en 2006 (MPO, 2008d). Pendant la saison de reproduction, le rassemblement des otaries aux quatre sites de reproduction canadiens rend la population de la Colombie-Britannique vulnérable aux perturbations humaines et aux événements catastrophiques, deux facteurs qui peuvent affecter une proportion importante de la population totale.
Alors que les facteurs limitatifs sont des processus naturels qui tendent à restreindre la taille d’une population ou à ralentir sa croissance, les menaces sont, de leur côté, les activités (tant naturelles qu’anthropiques) qui ont causé, qui causent ou qui peuvent causer des dommages, des mortalités ou des changements comportementaux chez une espèce en péril ou, encore, la destruction, la dégradation ou la détérioration de son habitat à un point tel que des effets seront ressentis par la population. La population d’otaries de Steller est limitée par des processus ascendants qui affectent la disponibilité et l’accessibilité des proies ainsi que par des processus descendants qui affectent les taux de prédation. Les menaces les plus importantes relevées pour l’otarie de Steller sont la concurrence exercée par les pêches et la variabilité environnementale qui limitent la disponibilité des proies. Par ailleurs, les déversements toxiques et la contamination chimique chronique sont modérément préoccupants au chapitre des impacts à long terme sur la santé de la population. D’autres recherches contribueront à clarifier l’importance de ces menaces.
L’otarie de Steller du Canada est inscrite à la liste de la Loi sur les espèces en péril en tant qu’« espèce préoccupante », ce qui signifie qu’elle est considérée comme une espèce sauvage qui pourrait devenir menacée ou en voie de disparition en raison d’une combinaison de caractéristiques biologiques et de menaces. Le présent plan de gestion a deux buts. Le premier est de faire en sorte que les menaces anthropiques d’origine canadienne ne provoquent pas un déclin insoutenable de la population ou une contraction de l’aire de répartition ou du nombre de sites de reproduction que l’on observe actuellement au Canada. Le second but du présent plan est d’appuyer les activités de recherche et de surveillance menées à l’égard des otaries de Steller en C.-B. qui permettent d’améliorer nos connaissances globales sur les populations de l’est du Pacifique, et de contribuer à ces activités. Six mesures hautement prioritaires concernant l’atténuation des menaces les plus préoccupantes (tableau 3) et 20 mesures en cours pour la conservation et la compréhension de l’espèce ont aussi été relevées. Six autres mesures sont indiquées concernant l’identification de nouveaux efforts utiles pour la gestion de la population. La participation des parties intéressées aux mesures précisées est la bienvenue et contribuera à la mise en œuvre du présent plan et à la conservation de l’espèce au Canada. La synchronisation des activités recommandées pour la protection, la gestion et la recherche facilitera l’adoption d’une approche plurispécifique axée sur la conservation des mammifères marins en C.-B. et permettra une utilisation efficace des ressources disponibles.
- PRÉFACE
- AUTORITÉS ET ORGANISMES RESPONSABLES
- AUTEURS
- REMERCIEMENTS
- ÉVALUATION ENVIRONNEMENTALE STRATÉGIQUE
- SOMMAIRE
- 1. RENSEIGNEMENTS SUR L’ESPÈCE
- 1.1. Information du COSEPAC sur l’évaluation de l’espèce
- 1.2. Description et biologie
- 1.3. Populations et aire de répartition
- 1.3.1. Répartition mondiale
- 1.3.2. Répartition canadienne
- 1.3.3. Tendances relatives à la population mondiale
- 1.3.4. Tendances relatives à la population canadienne
- 1.4. Besoins de l’otarie de Steller
- 1.4.1. Besoins en matière d’habitat et besoins biologiques
- 1.4.2. Rôle écologique
- 1.4.3. Facteurs limitatifs
- 1.5. Menaces
- 1.5.1. Classification des menaces
- 1.5.2. Description des menaces
- 1.5.3. Effets cumulatifs ou synergiques des menaces ou des facteurs limitatifs
- 1.6. Mesures déjà prises ou en cours
- 1.6.1. Gestion
- 1.6.2. Application de la réglementation
- 1.6.3. Évaluation des populations
- 1.7. Lacunes dans les connaissances
- 2. Gestion
- 2.1. But
- 2.2. Objectifs
- 2.3. Mesures
- 2.3.1. Protection
- 2.3.2. Gestion
- 2.3.3. Recherche sur la biologie de l’otarie de Steller
- 2.3.4. Recherche pour clarifier les menaces relevées
- 2.3.5. Suivi de l’état de la population
- 2.3.6. Vulgarisation et communication
- 3. Calendrier de mise en œuvre proposé
- 4. PLANS CONNEXES
- 5. RÉFÉRENCES
- 6. ANNEXE I
- 7. ANNEXE II
- 8. ANNEXE III : Collaboration et consultation
Figure 1. Aire de répartition mondiale de l’otarie de Steller. Les flèches indiquent les roqueries, et les zones ombrées, l’aire de répartition approximative des individus qui ne se reproduisent pas. La ligne pointillée illustre la séparation entre les stocks de l’Asie, de l’Est et de l’Ouest (adaptée d’une figure de Loughlin, 1997, et de Sease et al., 1999).
Figure 2. Emplacements géographiques des roqueries des otaries de Steller (l), échoueries occupées durant toute l’année (¡) et principales échoueries d’hiver (▲) en Colombie-Britannique. On peut également voir la roquerie principale située sur l’île Forrester, en Alaska. Mise à jour de Bigg (1985), d’après des travaux d’Olesiuk (données non publiées).
Figure 3. Tendances historiques concernant le nombre total d’otaries de Steller (petits, juvéniles et adultes) aux roqueries de la C.-B. (▲▬▲), à l’île Forrester, en Alaska (●▬●), et aux autres nouvelles roqueries du sud-est de l’Alaska (■▬■). La mince ligne bleue illustre la répartition des animaux entre les principales aires de reproduction de la C.-B. (adapté d’une figure de Bigg, 1985, et d’Olesiuk et al., 2008).
Figure 4. Nombre total déclaré d’otaries de Steller abattues en C.-B. dans le cadre des programmes d’abattage et des prélèvements commerciaux de 1913 à 1970. Les données ont été groupées et additionnées en périodes de cinq ans et sont représentées par des codes de couleur selon l’aire de reproduction principale. Une comparaison avec la figure 3 illustre l’impact qu’ont eu ces campagnes d’abattage et ces prélèvements sur les populations (données tirées de Bigg, 1984).
Tableau 1. Résumé de la classification des menaces et du potentiel d’atténuation des menaces relevées pour l’otarie de Steller, population de l’est du Pacifique. Le potentiel d’atténuation renvoie à la probabilité qu’une mesure (future ou actuelle) puisse atténuer les effets négatifs sur la population ou empêcher leur occurrence. La présente évaluation se veut un reflet à jour de l’état des menaces pesant sur la population; à ce titre, le classement de l’importance de la menace peut changer au fil du temps. L’astérisque (*) signifie que la menace est naturellement présente dans la population (c.-à-d. qu’il s’agit d’un facteur limitatif dont les effets peuvent être amplifiés par l’activité humaine).
Tableau 2. Les mesures de gestion indiquées dans le présent plan doivent être mises en œuvre, lorsque cela est approprié, en partenariat avec les organismes suivants.
Tableau 3. Proposition de calendrier de mise en œuvre
Tableau 4. Détails sur les termes utilisés pour l’évaluation des menaces pesant sur la population d’otaries de Steller de l’est du Pacifique.
Tableau 5. Polluants organiques persistants qui peuvent poser un risque pour l’otarie de Steller. Ce tableau provient de la version finale du programme de rétablissement des épaulards résidents au Canada (MPO, 2008).
Date de l’évaluation : Juillet 2005 Nom commun (population) : Otarie de Steller Désignation selon le COSEPAC : Espèce préoccupante Justification de la désignation : Il n’existe que trois1 sites de reproduction en Colombie-Britannique. Bien que la population augmente, cette espèce est vulnérable aux perturbations par les humains lorsqu’elle est sur terre. Parmi les menaces, il y a la possibilité de déversements graves d’hydrocarbures. D’autres populations, au nord et à l’ouest de la Colombie-Britannique, connaissent des déclins inexpliqués. Historique de la désignation du COSEPAC : Espèce désignée « non en péril » en avril 1987. Réexamen du statut et désignation de l’espèce comme étant « préoccupante » en novembre 2003. 1Depuis la désignation faite par le COSEPAC en 2003, un quatrième emplacement a été reclassé en tant que site de reproduction. |
L’otarie de Steller est le plus grand représentant des phoques à oreilles (ou otaries) et des otaries à fourrure (ordre des carnivores, superfamille des pinnipèdes, famille des Otariidés; Rice, 1998). Au Canada, elle porte aussi le nom de lion de mer de Steller. Certaines Premières nations l’appellent également « tukuk ou tukukw » (« tukwašt » signifiant « otarie séchée ») dans les dialectes nuu-chah-nulth du bassin de Barkley (BSDWG, 2004); en kwakiutl, les otaries sont appelées « tl’íx7en » (Grubb, 1977); dans la langue des Haïdas, elles sont appelées « kíit ou kíidaay » (Lawrence, 1977). Son nom scientifique, Eumetopias jubatus, signifie « qui a un large front et une crinière » et fait référence au collier proéminent de poils drus que portent les mâles adultes autour du cou et de la poitrine, d’où le nom de « lion de mer » que l’on donne parfois à l’espèce.
L’otarie de Steller présente un important dimorphisme sexuel, les mâles adultes atteignant une longueur de plus de 3 m et pesant entre 400 et 800 kg, bien que les plus gros d’entre eux puissent dépasser les 1 100 kg au début de la période de reproduction. Les mâles adultes développent un cou et une poitrine massifs et musclés, une tête robuste et un museau plus aplati que celui des femelles. Les femelles adultes sont nettement plus petites et atteignent en moyenne 2,2 m et entre 200 et 300 kg (Mathisen et al., 1962; Thorsteinson et Lensink, 1962; Orr et Poulter, 1967; Winship et al., 2001).
La fourrure des juvéniles et des adultes, lorsqu’elle est sèche, a une coloration variant du jaune pâle au brun clair et est plus sombre (brun chocolat) sur la face ventrale et près des nageoires, lesquelles sont noires et dépourvues de poils. Mouillée, cette fourrure peut apparaître d’un blanc grisâtre. À la naissance, les petits portent un lanugo épais d’un brun noirâtre jusqu’à la mue, qui se produit vers l’âge de 3 à 6 mois. L’otarie de Steller a un pelage fait de poils rudes et courts et subit une mue annuelle entre la fin de juin et le début de décembre, selon les classes d’âge (Scheffer, 1964). Les petits naissent avec un lanugo brun noirâtre épais et muent entre l’âge de 3 à 6 mois.
Toutes les otaries de Steller sont remarquablement agiles sur terre en raison de leur capacité de tourner leurs nageoires postérieures vers l’avant et de s’appuyer sur leurs nageoires antérieures. Les otaries de Steller peuvent grimper sur des rochers escarpés et sont souvent observées à de nombreux mètres au-dessus de la surface de la mer. Sur terre, elles ont tendance à être très grégaires et se rassemblent en groupes compacts dans des colonies de reproduction denses (roqueries) ou sur des échoueries ne servant pas à la reproduction (Schusterman, 1981; Loughlin et al., 1987). Bien que les otaries de Steller fréquentent habituellement des échoueries de façon régulière, elles passent parfois de nombreux jours ou plusieurs semaines en mer sans venir à terre (Olesiuk et Jeffries, données non publiées) et peuvent dormir dans l’eau, habituellement dans des groupes que l’on appelle des « radeaux ».
Les colonies reproductrices (appelées roqueries) sont peuplées d’otaries ayant atteint la maturité sexuelle (ainsi que de quelques jeunes qui dépendent de leur mère) pendant les mois d’été. L’otarie de Steller étant polygyne, les mâles exercent une compétition pour établir leur territoire et avoir accès aux femelles; le ratio entre les femelles et les mâles territoriaux est en général d’environ 10-15 : 1 (Gisiner, 1985; Merrick, 1987). Les mâles qui ont conquis un territoire gardent d’ordinaire celui-ci pendant 40 jours en moyenne (entre 20 et 68 jours), période durant laquelle ils ne s’alimentent pas (Gentry, 1970). La plupart des mâles territoriaux sont âgés de 9 à 13 ans (Thorsteinson et Lensink, 1962) et peuvent occuper un territoire pendant plusieurs années de suite (entre un an et sept ans) (Gisiner, 1985).
Les femelles ovulent pour la première fois entre l’âge de 3 et 6 ans. Elles commencent à revenir aux roqueries à la fin de mai et donnent naissance à un seul petit quelques jours après leur arrivée; la plupart des petits sont nés au début de juillet (Gentry, 1970; Edie, 1977; Bigg, 1985). Les mères allaitent leur petit sur le rivage pendant environ une semaine, puis amorcent des sorties régulières pour s’alimenter, passant un jour en mer et le suivant sur le rivage, et ainsi de suite (Swain, 1996). Au bout de quatre semaines environ, les petits peuvent nager en pleine mer, et les mères commencent à les amener aux échoueries adjacentes. Certains petits sont encore allaités lorsqu’ils entament leur troisième année, bien que la plupart soit sevrés au cours de leur première ou de leur deuxième année de vie. Certaines mères peuvent également allaiter un nouveau-né et un jeune d’un an simultanément (Sandegren, 1970; Hood et Ono, 1997; Milette et Trites, 2003).
L’utilisation des roqueries décline vers la fin d’août et demeure minimale de janvier à avril, quoique certains individus continuent à les utiliser toute l’année en tant qu’échoueries (Bigg, 1985). À l’extérieur de la saison de reproduction estivale, l’otarie de Steller utilise des échoueries pendant toute l’année ainsi que des échoueries d’hiver qui peuvent se situer à des distances considérables des roqueries. Les femelles et leur petit peuvent demeurer à une même échouerie ou passer d’une échouerie à l’autre (Shusterman, 1981; Loughlin et al., 1987).
L’otarie de Steller vit dans les eaux côtières fraîches tempérées et subarctiques du Pacifique Nord, depuis les îles Channel, au large du sud de la côte californienne, jusqu’au détroit de Béring au nord, et vers le sud-ouest le long de la côte asiatique jusqu’à l’île de Hokkaido, au Japon (figure 1; Kenyon et Rice, 1961; Loughlin et al., 1984; Loughlin et al., 1992). À l’heure actuelle, ces animaux mettent bas dans 61 roqueries et se reposent à plus de 300 échoueries dans cette aire de répartition. L’otarie de Steller n’est pas une espèce migratrice, mais les individus peuvent se disperser jusqu’à une distance considérable des sites de reproduction (Fisher, 1981; Calkins et Pitcher, 1982; Loughlin, 1997; Raum-Suryan et al., 2002).
À l’échelle mondiale, on distingue au moins deux populations d’otaries de Steller sur la base de la différenciation génétique de l’ADN mitochondrial (qui reflète la lignée maternelle) : la population de l’Est (de la Californie au sud-est de l’Alaska) et la population de l’Ouest (golfe d’Alaska, mer de Béring, îles Aléoutiennes et Russie) (Bickham et al., 1996). Une analyse phylogéographique qui tient compte de paramètres auxiliaires comme les tendances démographiques, la répartition, les déplacements et la morphologie conforte la distinction entre les deux populations en Amérique du Nord (York et al., 1996; Loughlin, 1997). Cependant, des échantillons génétiques prélevés récemment révèlent que deux nouvelles roqueries près de l’extrémité ouest de l’aire de répartition de la population de l’Est pourraient avoir été colonisées par un mélange d’individus des stocks de l’Ouest et de l’Est (O’Corry-Crowe et al., 2005). Les otaries de Steller femelles vivant dans les eaux asiatiques (péninsule du Kamchatka, Kouriles et mer d’Okhotsk) semblent être distinctes sur le plan génétique des autres populations (Baker et al., 2005), mais on observe un flux génétique supérieur chez les mâles (Hoffman et al., 2006). On dispose également de certaines preuves génétiques de ségrégation géographique entre les composants du plateau (golfe et péninsule de l’Alaska) et océaniques (Aléoutiennes) de la population de l’Ouest (O’Corry-Crowe, 2007).
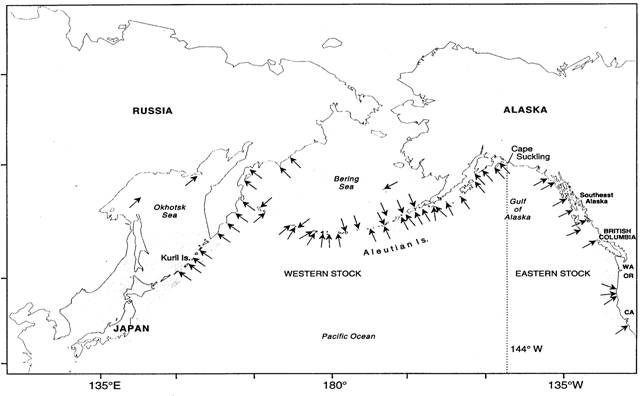
Figure 1. Aire de répartition mondiale de l’otarie de Steller. Les flèches indiquent les roqueries, et les zones ombrées, l’aire de répartition approximative des individus qui ne se reproduisent pas. La ligne pointillée illustre la séparation entre les stocks de l’Asie, de l’Est et de l’Ouest (adaptée d’une figure de Loughlin, 1997, et de Sease et al., 1999).
Au Canada, l’otarie de Steller n’est présente qu’en Colombie-Britannique et fait partie de la population de l’Est (Bickham, 2000). On recense quatre aires de reproduction principales en C.-B. : la première aux îles Scott, au large de la pointe nord-ouest de l’île de Vancouver, les roqueries étant situées sur l’île Triangle et sur les petits îlots au large des îles Beresford et Sartine; la deuxième à cap St. James, au large de la pointe sud des îles de la Reine-Charlotte, les roqueries étant situées dans les îles Kerouard; la troisième au large de l’île Banks, dans le nord de la côte continentale, les roqueries étant situées sur les rochers North Danger (figure 2).
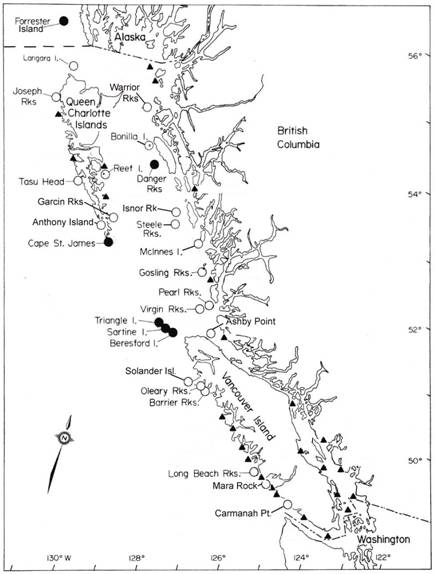
Figure 2. Emplacements géographiques des roqueries des otaries de Steller (l), échoueries occupées durant toute l’année (¡) et principales échoueries d’hiver (▲) en Colombie-Britannique. On peut également voir la roquerie principale située sur l’île Forrester, en Alaska. Mise à jour de Bigg (1985), d’après des travaux d’Olesiuk (données non publiées).
Une quatrième aire de reproduction a déjà existé au large de la côte continentale centrale, dans le groupe Sea Otter, les roqueries étant situées sur les rochers Virgin, Pearl et (peut-être) Watch, mais cette zone de reproduction a été abandonnée à la suite des programmes d’abattage intenses des prédateurs exécutés dans les années 1920 et 1930; elle a été, par la suite, utilisée en tant qu’échouerie par des individus ne se reproduisant pas (Bigg, 1985). Cependant, un nombre croissant de petits sont nés sur les rochers Virgin et Pearl au cours des dernières années et, selon le relevé de l’abondance effectué en 2006, on a reclassé le site en tant que roquerie. Outre les quatre sites de reproduction, on recense environ 23 échoueries, réparties principalement sur le front océanique de la côte, qui sont utilisées de façon continue toute l’année, ainsi que de nombreux sites d’hiver utilisés sur une base saisonnière ou irrégulière.
La répartition des otaries de Steller au large est mal définie. En général, la plupart des otaries de Steller semblent se nourrir à moins de 60 km du rivage en été et peuvent s’éloigner jusqu’à 200 km du rivage en hiver (Kenyon et Rice, 1961; Merrick et Loughlin, 1997). L’espèce semble se nourrir au-dessus de la plate-forme continentale et le long du bord de celle-ci (Kajimura et Loughlin, 1988). Des otaries de Steller capturées en C.-B. et munies d’émetteurs satellites ont effectué des déplacements importants le long de la côte de la C.-B., mais se sont rarement aventurées à plus de 50 km de la côte (Olesiuk et Jeffries, données non publiées).
On croit que, de la fin des années 1950 aux années 1970, l’abondance globale de l’otarie de Steller dans le Pacifique Nord (dans toute l’aire de répartition, de la Californie au Japon) a été stable, se situant aux alentours de 250 000 à 300 000 individus (Kenyon et Rice, 1961; Loughlin et al., 1984). La population a décliné par la suite pour atteindre environ 116 000 individus en 1989, 97 500 entre 1994 et 1995, et 95 000 entre 1999 et 2002.
La chute de la population totale d’otaries de Steller est attribuable au déclin de l’espèce dans la portion ouest de son aire de répartition. Historiquement, la population de l’Ouest (de la Russie au golfe d’Alaska) était beaucoup plus importante que la population de l’Est (du sud-est de l’Alaska à la Californie); entre les années 1950 et les années 1970, elle représentait environ 90 % de la population totale (Kenyon et Rice, 1961; Loughlin et al., 1984; Trites et Larkin, 1996). Le déclin semble avoir débuté dans la partie est des Aléoutiennes vers 1965 pour se propager au reste de l’archipel et au golfe d’Alaska un peu avant 1980. La population a chuté abruptement pendant les années 1980, puis à un rythme beaucoup plus lent après 1990 (York et al., 1996). Entre 1999 et 2002, on estimait la population de l’Ouest à environ 50 000 individus (Burkanov, 2000; Sease et Stinchcomb, 2003), ce qui représente un déclin d’environ 80 % comparativement aux populations observées des années 1950 aux années 1970. Le déclin précipité de la population de l’Ouest a fait de l’otarie de Steller l’un des mammifères marins les plus étudiés dans le monde (voir NMFS, 1992, 2007; Loughlin, 1998; Hunter et Trites, 2001; Dalton, 2005).
En raison de la mise en vigueur du programme d’abattage des prédateurs en C.-B., dans l’État de Washington, en Oregon (Rowley, 1929) et en Californie (Pearson et Verts, 1970) au cours de la majeure partie du XXe siècle (1900-1970), la population d’otaries de Steller de l’Est était fortement décimée au moment où elle est devenue une espèce protégée en vertu de la Loi sur les pêches du Canada, en 1970, et de la Marine Mammal Protection Act des États-Unis, en 1972.
L’otarie de Steller ne se reproduisait pas dans le sud-est de l’Alaska au début des années 1900 et n’a donc pas fait l’objet de programmes d’abattage majeurs. Les premières roqueries ont vu le jour dans le sud-est de l’Alaska au cours des années 1930 ou 1940, alors que l’abattage intense avait débuté dans les parties sud de l’aire de répartition de l’espèce, et cette dernière semble avoir proliféré dans les années 1950 et 1960, même si les populations reproductrices étaient réduites en C.-B. Pendant que les programmes d’abattage des prédateurs étaient en cours, la dispersion de femelles reproductrices provenant des roqueries de la C.-B. peut avoir contribué à l’expansion rapide de nouvelles roqueries établies dans le sud-est de l’Alaska (Calkins et al., 1999; Pitcher et al., 2003, 2007), et les profils de dispersion semblent avoir évolué au fil du temps. L’île Forrester a été le théâtre de la majeure partie de la croissance de la population jusqu’au début des années 1980, mais le taux de croissance y a par la suite ralenti, et de nouvelles roqueries établies dans le centre nord du sud-est de l’Alaska ainsi que d’autres roqueries établies en C.-B. comptent maintenant pour la majeure partie de la croissance récente (Pitcher et al., 2007).
Contrairement à la population de l’Ouest, la population de l’Est s’est accrue ces dernières années (MMO, 2008d; Olesiuk, données non publiées; Calkins et al., 1999; Pitcher et al., 2003, 2007). En 2002, la population de l’Est a été estimée à environ 46 000-58 000 individus (Pitcher et al., 2007), comparativement à 45 000 individus dans l’ouest de l’Alaska (Angliss et Outlaw, 2007) et à 16 000 individus en Asie (Burkanov et Loughlin, 2007).
Les dénombrements historiques de roqueries d’otaries de Steller en C.-B. remontent au début des années 1900 et fournissent un indice de la taille de la population reproductrice canadienne (Bigg, 1984, 1985; MPO, 2008d). On estime qu’environ 14 000 individus (de tous les âges, y compris les petits) étaient présents sur les roqueries entre 1913 et 1919 (figure 3), époque antérieure aux grandes campagnes d’abattage. Les otaries de Steller vivant dans les eaux canadiennes et avoisinantes ont fait l’objet de programmes d’abattage des prédateurs pendant la majeure partie du XXe siècle.
Avant que l’espèce ne devienne protégée, dans les années 1970, le gouvernement canadien a mené des campagnes d’abattage intensives des otaries de Steller aux roqueries afin de protéger les pêches au saumon (figure 4). Des prélèvements commerciaux limités ont également eu lieu pour la production de cuir et d’aliments pour les visons. Le nombre d’individus aux roqueries de la C.-B. avait été réduit à 4 550 en 1961, et à 3 390 (y compris 940 petits) au moment où le premier relevé aérien a été effectué, en 1971. Ainsi, la population d’otaries de Steller de la C.-B. a été réduite à environ le quart de son effectif historique par les programmes d’abattage des prédateurs et les prélèvements commerciaux (Bigg, 1985; MPO, 2008d).
Les évaluations de l’abondance, de la répartition et de l’état des stocks d’otaries de Steller en C.-B. ont débuté dans les années 1970 et 1980 (Bigg, 1984, 1985, 1988) et se sont poursuivies jusqu’à ce jour (MPO, 2008d; Olesiuk et al., données non publiées). Des relevés aériens couvrant toute la province ont également été menés en C.-B. à des intervalles de quatre à cinq ans depuis que l’espèce est protégée, c’est-à-dire depuis 1970. Les relevés sont menés pendant une brève période se situant entre la fin de juin et le début de juillet, alors que la majeure partie des petits sont nés, mais sont encore trop jeunes pour avoir commencé à se disperser hors des roqueries (Bigg, 1985; MPO, 2008d).
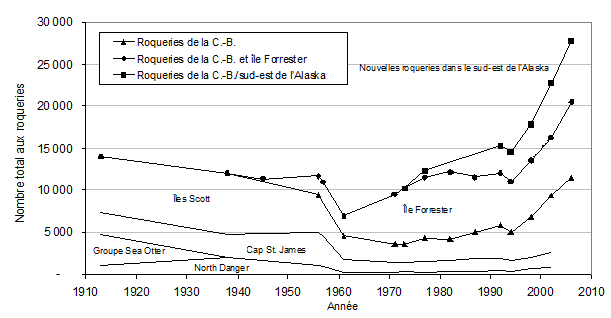
Figure 3. Tendances historiques concernant le nombre total d’otaries de Steller (petits, juvéniles et adultes) aux roqueries de la C.-B. (▲▬▲), à l’île Forrester, en Alaska (●▬●), et aux autres nouvelles roqueries du sud-est de l’Alaska (■▬■). La mince ligne bleue illustre la répartition des animaux entre les principales aires de reproduction de la C.-B. (adapté d’une figure de Bigg, 1985, et d’Olesiuk et al., 2008).
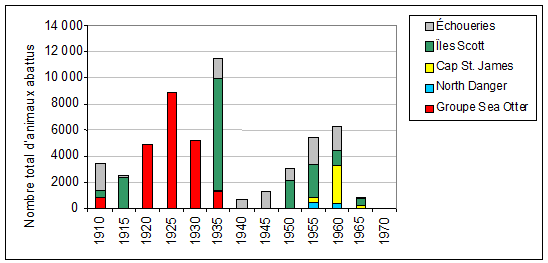
Figure 4. Nombre total déclaré d’otaries de Steller abattues en C.-B. dans le cadre des programmes d’abattage et des prélèvements commerciaux de 1913 à 1970. Les données ont été groupées et additionnées en périodes de cinq ans et sont représentées par des codes de couleur selon l’aire de reproduction principale. Une comparaison avec la figure 3 illustre l’impact qu’ont eu ces campagnes d’abattage et ces prélèvements sur les populations (données tirées de Bigg, 1984).
Les relevés aériens effectués en C.-B. révèlent que le nombre de petits et d’individus autres que des petits présents sur les roqueries s’est accru à un rythme moyen de 3,5 % et de 3,9 % par année respectivement, ce qui a fait en sorte que la taille de la population reproductrice a triplé depuis le début des années 1970 (figure 5) (MPO, 2008d). Des tendances similaires ont été observées aux roqueries avoisinantes en Oregon, au sud, et dans le sud-est de l’Alaska, au nord (figure 5) ; ces deux régions combinées recèlent 90 % de la population de l’Est (Brown et Reimer, 1992; Calkins et al., 1999; Pitcher et al., 2007). Cela révèle que l’augmentation observée au Canada représente une croissance réelle de la population et non uniquement un déplacement local de la répartition des individus (Pitcher et al., 2007).
Selon l’estimation pour la production de petits dérivée du dernier relevé mené sur l’ensemble de l’aire de répartition en 2002, la taille totale de la population de l’Est oscillerait entre 46 000 et 58 000 individus, dont un tiers (33 % de petits et 34 % d’individus autres que des petits) fréquenterait les eaux canadiennes. On a estimé que l’abondance dans les eaux canadiennes était de 20 000 à 28 000 individus selon le relevé le plus récent mené en 2006 (MPO, 2008d).
Les otaries de Steller n’ont pas les mêmes besoins sur terre, lorsqu’elles sortent de l’eau pour se reposer et se reproduire, qu’en mer, où elles se déplacent et s’alimentent. L’espèce est grégaire lorsqu’elle est sur terre, les animaux se rassemblant aux roqueries et aux échoueries traditionnelles, dont certaines sont utilisées depuis plus d’un siècle. L’otarie de Steller s’alimente d’une multitude de proies, principalement de poissons de petite et de moyenne taille vivant en bancs, et l’habitat d’alimentation varie selon la répartition et l’abondance des proies.
Habitat terrestre
Les sites terrestres fréquentés par l’otarie de Steller en Colombie-Britannique se divisent de façon générale en trois catégories distinctes : 1) les roqueries, où les otaries se rassemblent de mai à août pour s’accoupler, mettre bas et allaiter les petits; 2) les échoueries permanentes, généralement occupées toute l’année; 3) les échoueries hivernales, qui sont occupées moins régulièrement, principalement en dehors de la saison de reproduction (Bigg, 1985). En général, les roqueries sont avoisinées d’échoueries, lesquelles sont principalement occupées par des mâles qui ne s’accouplent pas et des juvéniles, pendant la période de reproduction. Dans la plupart des cas, des individus continuent d’utiliser les roqueries comme échoueries toute l’année, bien qu’en nombre très réduit.
L’otarie de Steller affiche une grande fidélité aux sites de reproduction et de mise bas. Des études portant sur des individus marqués indiquent que la plupart des femelles tendent à retourner à la roquerie où elles sont nées et qu’elles retournent fidèlement à la même roquerie chaque année (Raum-Suryan et al., 2002). Les trois principales colonies reproductrices de la C.-B. étaient toutes bien établies lorsque le premier relevé visant l’espèce a été mené en 1970 (Newcombe et Newcombe, 1914), et celles-ci ont été utilisées de façon continue malgré les perturbations causées par les programmes d’abattage des prédateurs et les prélèvements commerciaux (Pike et Maxwell, 1958; Bigg, 1985; Olesiuk, données non publiées). Une quatrième aire de reproduction, située dans le groupe Sea Otter, a été abandonnée à la suite des programmes d’abattage des prédateurs, même si elle a été utilisée en tant qu’échouerie par des individus ne se reproduisant pas. La mise bas a recommencé à ce site et, d’après les dénombrements des petits effectués pendant le relevé de 2006, on a récemment reclassé cet emplacement en tant que roquerie.

Même si les otaries de Steller retournent en général à leur roquerie natale pour se reproduire, il peut également y avoir certains échanges avec des roqueries avoisinantes (Calkins et Pitcher, 1982, 1996). Sur les 31 femelles marquées alors qu’elles étaient petites sur l’île Forrester, plusieurs ont été observées en train de mettre bas à d’autres roqueries, y compris à celles situées à 400 km, au cap St. James (Raum-Suryan et Pitcher, 2000; Raum-Suryan et al., 2002).
Les habitats terrestres utilisés par les otaries de Steller comprennent certains des affleurements rocheux dénudés les plus isolés du Pacifique Nord. Les échoueries sont d’ordinaire situées dans des régions qui affichent des courants relativement forts, une salinité élevée, de faibles températures de surface et des eaux peu profondes, ce qui témoigne vraisemblablement d’une forte productivité océanique et, de ce fait, de la présence d’aires d’alimentation optimales (Ban et al., données non publiées). Les caractéristiques essentielles des échoueries semblent inclure la présence d’un terrain relativement plat, des lieux accessibles, une protection contre les lames et les vagues ainsi que l’absence de prédateurs terrestres tels que les ours et les loups (Edie, 1977). Les otaries utilisent des aires protégées pendant les tempêtes et les aires humides durant les périodes de très grande chaleur (Edie, 1977). L’accès à des terrains élevés est également important pour la mise bas, même si les individus plus âgés qui sont capables d’aller en mer utilisent les zones moins élevées et plus exposées. Les barres rocheuses constituent le substrat de reproduction de prédilection dans les roqueries de la C.-B., même si un nombre croissant d’individus ont commencé dernièrement à se reproduire sur des plages de gravier le long du côté est (sous le vent) de l’île Triangle (Olesiuk, données non publiées).
Les 23 échoueries permanentes de Colombie-Britannique, situées en général sur le front océanique le long de la côte ouest, comprennent des affleurements et des îlots rocheux. Environ la moitié d’entre elles ont été signalées en 1913, lors des premiers relevés (Newcombe et Newcombe, 1914), et environ le quart semblent avoir été colonisées après le début des années 1970, soit depuis les premiers relevés aériens. Les échoueries permanentes sont largement réparties en C.-B. et offrent aux otaries un habitat pour se reposer tout le long de la côte océanique. Les otaries de Steller peuvent également demeurer dans l’eau pendant les tempêtes ou les périodes de fortes lames, alors que les échoueries sont balayées par les vagues, ou lorsqu’elles se trouvent près de concentrations de proies sans disposer d’échoueries appropriées à proximité; ce comportement d’« agrégation » est observé fréquemment (Kenyon et Rice, 1961; Olesiuk et Bigg, 1988).
La plupart des échoueries hivernales sont situées dans des zones abritées comme les détroits de Georgia, de Juan de Fuca et de la Reine-Charlotte. Outre les substrats naturels, on trouve dans les échoueries hivernales des estacades, des flots, des jetées et des quais. Dans la partie sud de la Colombie-Britannique, les otaries de Steller partagent souvent leurs échoueries hivernales avec des otaries de Californie (Zalophus californianus) mâles subadultes et adultes (Hancock, 1970; Brenton, 1977; Bigg, 1985).
On a identifié plus de 50 espèces de poissons et d’invertébrés dans le régime alimentaire de l’otarie de Steller (Wilke et Kenyon, 1952; Pike, 1958; Spalding, 1964; Pitcher, 1981; Kastelein et al., 1990; Sinclair et Zeppelin, 2002). En Colombie-Britannique, les proies de prédilection de l’otarie de Steller semblent être des poissons de petite et de moyenne taille vivant en bancs, dont le hareng, la merluche, le lançon, le saumon, l’aiguillat, l’eulakane et la sardine, ainsi que des poissons de fond comme le sébaste, les poissons plats et les raies (Pike, 1958; Spalding, 1964; Olesiuk et Bigg, 1988, Trites et Olesiuk, données non publiées). Outre les poissons, les otaries s’alimentent parfois de pieuvres et de calmars, mais l’importance de ces proies dans le régime alimentaire des otaries a vraisemblablement été exagérée dans les études antérieures du fait que les becs des céphalopodes peuvent s’accumuler sur de longues périodes dans l’estomac des otaries (Bigg et Fawcett, 1985). Les crabes, les moules, les myes et d’autres invertébrés sont parfois récupérés dans les contenus stomacaux et les excréments, mais il peut s’agir de proies secondaires qui ont été consommées par les proies dont les otaries se sont nourries. On a également observé des otaries de Steller s’attaquer à des goélands (O’Daniel et Schneeweis, 1992) et à d’autres pinnipèdes, y compris des petits d’otaries à fourrure (Gentry et Johnson, 1981) et de phoques communs (Pitcher et Fay, 1982; E. Mathews, Université de l’Alaska, Juneau AK, comm. pers.). La prédation visant d’autres pinnipèdes semble plutôt rare, mais pourrait être importante à certains endroits.
Les besoins des otaries de Steller en matière de proies varient d’une saison à l’autre selon l’âge, le sexe, le type de proies présentes et leur qualité (Perez,1994; Rosen et Trites, 1999, 2000b,c). Des modèles bioénergétiques indiquent que les besoins alimentaires quotidiens des otaries de Steller sauvages sont d’environ 15 à 20 kg chez les femelles matures et de 30 à 35 kg chez les mâles matures (Winship et al., 2002). Dans le cas des femelles, ces besoins énergétiques quotidiens représentent environ 14 % de la masse corporelle d’une femelle d’un an et 7 % de la masse corporelle d’une femelle mature. La consommation moyenne au sein de la population du sud-est de l’Alaska a été estimée à 17 kg par individu et par jour (Winship et Trites, 2003). Les otaries qui consomment davantage de poissons à faible teneur en gras, tels les gadidés, doivent consommer beaucoup plus de proies que ceux qui s’alimentent de poissons plus riches en gras, comme le hareng (Trites et Donnelly, 2003; Winship et Trites, 2003).
On ne sait pas très bien comment les otaries de Steller exploitent leur habitat aquatique. En mer, on voit les otaries de Steller seules ou en groupes de plusieurs individus (Bonnell et al.1983). Les otaries qui se nourrissent de petits poissons vivant en bancs semblent s’alimenter en coopération, leurs groupes pouvant réunir jusqu’à 100 individus qui plongent et font surface en synchronie (Fiscus et Baines, 1966; Loughlin et al., 1983; Loughlin et DeLong, 1983; P. Olesiuk, obs. pers.). La recherche de nourriture semble se faire surtout la nuit, d’après la télémétrie par satellite (Loughlin et al., 1998; Loughlin et al., 2003) et la tendance qu’a l’animal à aller à terre le jour (Withrow, 1982; Higgins, 1984; Milette, 1999), mais peut varier d’une saison à l’autre selon le type de proies consommées (Olesiuk et Jeffries, données non publiées). Les otaries de Steller peuvent plonger à des profondeurs de 310 m et plus (Andrews, 1999), et demeurer sous l’eau pendant plus de huit minutes (Swain et Calkins, 1997); la plupart des plongées ont lieu entre 15 et 50 m de profondeur et durent entre 1,5 et 2,5 min (Merrick et Loughlin, 1997; Swain et Calkins, 1997; Loughlin et al., 1998; Andrews, 1999; Swain, 1999).
On observe généralement les individus à moins de 60 km de la terre, dans des eaux d’une profondeur inférieure à 400 m, mais ils peuvent s’aventurer à plusieurs centaines de kilomètres des côtes et au large de la plate-forme continentale (Kenyon et Rice, 1961; Merrick et Loughlin, 1997). Les études par télémétrie et par marquage ont démontré que les otaries sont très mobiles et qu’elles peuvent parcourir des milliers de kilomètres et utiliser de nombreuses échoueries au cours de périodes allant de quelques semaines à quelques mois (Merrick et Loughlin, 1997; Loughlin et al., 1997, 2003; Raum-Suryan et al., 2002). Des otaries de Steller capturées en C.-B. et marquées ont été par la suite suivies sur une distance allant jusqu’à 1 700 km, vers le nord en Alaska ou vers le sud, en Californie (Calkins, 1981; Fisher, 1981; Loughlin, comm. pers.; Olesiuk, données non publiées). Les otaries de Steller s’aventurent à l’occasion en eaux douces (Jameson et Kenyon, 1977; Roffe et Mate, 1984; Beach et al., 1985). En C.-B., les otaries sont observées à l’occasion en groupe sur une distance allant jusqu’à 35 km en amont (Olesiuk, données non publiées). Les otaries de Steller se rassemblent également dans les estuaires à l’automne pour s’alimenter de saumons prégénésiques et à l’embouchure du fleuve Fraser au printemps, lorsqu’a lieu la montaison de l’eulakane (Bigg, 1985; Bigg et al., 1990, Olesiuk, données non publiées). Il semblerait que les principales zones d’hivernage des otaries au large du sud de l’île de Vancouver varient en fonction de la répartition des harengs prégénésiques (P. Olesiuk, comm. pers.).
Les sorties d’alimentation ont tendance à être davantage localisées pendant la saison de reproduction estivale (< 20 km) qu’en toute autre période de l’année (60-160 km) (Bonnell et al., 1983; Merrick et Loughlin, 1997). Au cours de l’été, les femelles reproductrices doivent s’alimenter près des roqueries du fait qu’elles doivent revenir régulièrement pour s’occuper de leur petit. Par contre, les individus qui ne se reproduisent pas pendant l’été ainsi que l’ensemble des individus en dehors de la saison de reproduction (de septembre à mai) disposent de plus de latitude et se rendent vraisemblablement là où des poissons fourrages sont présents. La superficie de l’aire d’alimentation des individus immatures non reproducteurs semble se situer entre les superficies des aires d’alimentation estivale et hivernale des adultes (Merrick et Loughlin, 1997).
L’otarie de Steller est l’espèce la plus grande de la famille des Otariidés et est la seule qui réside en permanence et qui se reproduit dans les eaux canadiennes. L’espèce occupe un créneau intermédiaire entre l’aire de répartition côtière du phoque commun (Phoca vitulina), qui vit généralement dans des eaux plus protégées, et l’aire de répartition pélagique de l’otarie à fourrure du Nord (Callorhinus ursinus), qui vit généralement sur le plateau continental et le long du bord de ce même plateau. Le rôle des phoques et des otaries dans les écosystèmes marins complexes demeurant mal connu, il faut faire des études pour évaluer la contribution de l’otarie de Steller dans les écosystèmes complexes et de grande envergure (Beverton, 1985; Bowen, 1997; Merrick, 1997; Trites, 1997).
En raison des déclins récents observés en Alaska, les roqueries de la Colombie-Britannique situées aux îles Scott et au cap St. James figurent dorénavant aux deuxième et sixième rangs mondiaux pour ce qui est de l’importance des agrégations de reproducteurs. D’après la production totale de petits en 2002, la C.-B. soutient environ 16 % de la population mondiale et environ 33 % du stock de l’Est (une autre proportion de 31 % est présente dans le sud-est de l’Alaska, à moins de 50 km de la frontière canadienne). De façon générale, les otaries de Steller sont considérées comme un composant important de l’écosystème marin côtier et contribuent à l’industrie de l’écotourisme.
L’otarie de Steller est un prédateur marin situé en haut de la chaîne alimentaire. Dans le sud-est de l’Alaska, on estime que l’otarie de Steller consomme environ 140 millions de kilogrammes de poissons chaque année (Winship et Trites, 2003); si l’on suppose un régime alimentaire similaire, la population de la C.-B. en consommerait 110 millions de kilogrammes supplémentaires chaque année. En comparaison, les débarquements annuels totaux de la pêche commerciale en C.-B. se sont chiffrés à environ 185 millions de kilogrammes au cours de la dernière décennie (MPO, 2007). On ne dispose toujours pas des connaissances de base concernant les habitudes alimentaires saisonnières et régionales des otaries en C.-B., la majeure partie de l’information dont on dispose ayant été recueillie de façon anecdotique dans le cadre d’autres études. Comme les otaries de Steller sont en rétablissement après la mise en œuvre de programmes d’abattage des prédateurs et des prélèvements commerciaux, il est possible que les ressources en matière de proies constituent, en bout de ligne, une limite à l’accroissement des populations d’otaries, mais on ne sait pas précisément dans quelle mesure les otaries peuvent à leur tour limiter les populations des proies qu’elles consomment.
Les otaries de Steller sont également en concurrence avec d’autres prédateurs marins, y compris d’autres espèces de pinnipèdes, des baleines, des oiseaux de mer, des requins et des poissons plats (Livingston, 1991; Tamura et Ohsumi, 2000; Wespestad et al., 2000; NMFS, 2001; Gallucci et al., 2006). Au cours du siècle dernier, la concurrence avec l’otarie de Californie (Zalophus californianus) s’est accrue de façon marquée (Lowry et Maravilla-Chavez, 2005), ce qui pourrait avoir eu une incidence négative sur l’otarie de Steller. L’otarie de Californie semble avoir chassé l’otarie de Steller de ses roqueries traditionnelles dans les îles Channel, au large de la Californie, et a agrandi son aire de répartition (en dehors des périodes de reproduction) vers le nord jusqu’en C.-B. (Bigg, 1988; P. Olesiuk, comm. pers.) et, dans certains cas, aussi loin qu’en Alaska (Maniscalco et al., 2004). Les otaries de Californie qui migrent le long de la côte de l’Oregon semblent en avoir chassé les otaries de Steller (Mate, 1975); les deux espèces partagent souvent les mêmes échoueries d’hiver (P. Olesiuk, comm. pers.) et consomment les mêmes espèces-proies (Olesiuk et Bigg, 1988).
L’otarie de Steller représente une proie importante pour l’épaulard migrateur (Morton, 1990; Baird et Dill, 1995; Ford et al., 1998; Matkin et al., 2007; Wade et al., 2007), lequel peut s’alimenter de façon sélective de petits et de juvéniles (Barrett-Lennard et al., 1995). L’épaulard migrateur est inscrit à la liste de la Loi sur les espèces en péril (LEP) en tant qu’espèce menacée et peut être vulnérable aux fluctuations affichées par les proies qu’il consomme. En C.-B. et dans les eaux adjacentes, l’otarie de Steller figure au deuxième rang en importance parmi les proies des épaulards migrateurs (Ford et al., 1998), et un relevé mené dans l’ensemble du Pacifique Nord-Est a révélé que l’otarie de Steller était la sixième proie de l’épaulard migrateur la plus couramment observée (Wade et al., 2007). Les grands requins s’alimentent également d’otaries de Steller dans la partie sud de l’aire de répartition de l’espèce (Stroud, 1978; Ainley et al., 1981), et le requin nourrice constitue un prédateur potentiel des otaries en Alaska (Sigler et al., 2006).
L’otarie de Steller peut servir d’indicateur de l’état général des écosystèmes marins côtiers. L’espèce affiche une répartition étendue dans les eaux côtières, est longévive, se regroupe sur des roqueries où il est possible de procéder au recensement des populations reproductrices et occupe une position située près du haut de la chaîne alimentaire marine. Les déclins observés récemment dans la population de l’Ouest, en Alaska, sont maintenant largement considérés comme étant associés à des processus écosystémiques de plus grande envergure que l’on comprend mal; cela démontre que la capacité d’assurer un suivi des populations d’otaries de Steller dépasse de beaucoup notre compréhension des processus écologiques complexes qui régularisent les populations de ces prédateurs du haut de la chaîne. Comme les populations de la C.-B. et des eaux environnantes se sont maintenant rétablies et ont même dépassé les effectifs élevés qu’elles ont déjà affichés par le passé, les mécanismes de régularisation naturelle des populations peuvent devenir un facteur régissant l’état des populations d’otaries de Steller.
Les facteurs limitatifs sont les processus naturels qui limitent la taille ou la croissance des populations. L’otarie de Steller est une espèce relativement longévive qui se reproduit lentement. La productivité maximale de cette espèce n’a pas été déterminée, mais elle est vraisemblablement faible. Dans le cadre des évaluations des stocks américaines, un taux de croissance maximal théorique de 12 % est généralement utilisé pour les pinnipèdes. Même si un tel taux peut convenir pour les phocidés (p. ex. Olesiuk et al., 1990; Olesiuk, 1999; Bowen et al., 2003), les Otariidés affichent en général des taux de survie moins élevés, et les populations en expansion n’ont pas atteint ce degré de productivité. Par exemple, plusieurs populations décimées d’otaries de Californie ont maintenu des taux de croissance soutenue exponentiels d’environ 6,1 % (Lowry et Maravilla-Chavez, 2005), tandis que des populations d’otaries à fourrure du Nord ont affiché des taux de croissance de 8,6 % par année (York, comm. pers., cité dans Angliss et Outlaw, 2007). Les populations d’otaries de Steller qui se rétablissent le long de la côte ouest de l’Amérique du Nord ont affiché des taux de croissance soutenue d’environ 3,1 % par année et n’ont présenté aucun signe de dépendance à la densité alors que les populations augmentaient, mais on ne sait pas si cela représente le taux de croissance intrinsèque maximal pour cette espèce ou si certains facteurs de perturbation ont ralenti le rétablissement dans cette région au cours des 40 dernières années (Pitcher et al., 2007). Néanmoins, la productivité des otaries de Steller est probablement inférieure à celle d’autres pinnipèdes, ce qui les rend moins résilientes aux perturbations et aux contraintes et fait en sorte qu’il leur faut plus de temps pour se rétablir à la suite de tels impacts.
La faible productivité des otaries de Steller peut être la résultante d’un faible potentiel reproducteur doublé d’une mortalité élevée. La plupart des Otariidés, y compris les otaries de Californie et les otaries à fourrure du Nord, sèvrent leurs petits lorsque ceux-ci ne sont âgés que de quelques mois et ont tendance à se reproduire chaque année. Par contre, l’otarie de Steller continue souvent à allaiter ses petits au cours des deuxième et troisième années de vie (Pitcher et al., 2003). Comme les femelles en lactation sont moins susceptibles de mener leur fœtus à terme, la période prolongée de soins parentaux entraîne d’ordinaire des intervalles plus longs entre les naissances et, dans l’ensemble, réduit le rendement reproducteur (Pitcher et al., 1998).
La mortalité chez les petits de moins de 1 mois est élevée (Pike et Maxwell, 1958; Orr et Poulter, 1967). La mortalité chez les juvéniles est difficile à estimer en raison d’éventuels biais associés à l’échantillonnage, mais semble être élevée : environ 48 % des femelles et 26 % des mâles survivent jusqu’à l’âge de 3 ans (Calkins et Pitcher, 1982; York, 1994). Les taux de mortalité plus élevés chez les mâles entraînent un rapport entre les sexes progressivement biaisé en faveur des femelles. Les taux de mortalité sont de beaucoup inférieurs chez les adultes (~ 10-15 % par année pour les femelles et ~ 13-25 % pour les mâles). La principale cause de mortalité chez les petits est la noyade du fait que ceux-ci peuvent difficilement nager à cet âge et qu’ils ne peuvent ressortir de l’eau ou se déplacer dans les forts courants maréaux (Orr et Poulter, 1967; Edie, 1977). Les morsures, les écrasements ou les piétinements infligés par des individus plus âgés et l’abandon par leur mère ou la séparation d’avec celle-ci contribuent également à la mortalité chez les petits (Orr et Poulter, 1967; Gentry, 1970; Sandegren, 1970; Sandegren, 1976). Chez les mammifères marins, la dépendance à la densité est, en général, exprimée principalement dans les paramètres qui affectent les taux de reproduction, particulièrement chez les jeunes animaux (c.-à-d. l’âge à la première mise bas, les taux de fécondité et la survie chez les juvéniles) (Eberhardt, 1985; Fowler, 1987). La mortalité juvénile est le principal facteur responsable des déclins spectaculaires observés chez la population de l’Ouest dans les années 1980 (York, 1994), la natalité réduite chez les femelles et la survie de celles-ci jouant un rôle moins important (Holmes et York, 2003). Le déclin plus lent observé dans les années 1990 peut être attribuable à une amélioration de la survie chez les juvéniles et les adultes, malgré des taux de natalité qui semblent avoir continué à chuter (Holmes et York, 2003). Le ratio entre les petits et les autres individus fournit un indice des taux de naissance et de survie relatifs. Le ratio étonnamment élevé des autres individus par rapport aux petits constaté à partir des relevés menés le long de la côte ouest de l’Amérique du Nord porte à croire que la survie accrue des juvéniles (par opposition à une augmentation des taux de natalité) a été un facteur important dans l’augmentation de la population de l’Est (Pitcher et al., 2007).
Les facteurs qui, en bout de ligne, limitent les populations d’otaries de Steller et d’autres prédateurs marins peuvent être, de façon générale, catégorisés soit comme étant des processus ascendants régis par la disponibilité et la qualité des proies, soit comme des processus descendants régis par les prédateurs (y compris les mortalités anthropiques directes).
Disponibilité et qualité des proies
La disponibilité de proies appropriées limite en bout de ligne les populations d’otaries de Steller. Un changement dans la qualité des régimes alimentaires, lesquels sont passés de poissons gras (c.-à-d. de harengs) à des poissons à faible teneur en gras (c.-à-d. la goberge de l’Alaska), est en partie responsable du déclin des otaries de Steller dans le golfe d’Alaska et aux Aléoutiennes (Alverson, 1992; Alaska Sea Grant, 1993; DeMaster et Atkinson, 2002; Trites et Donnelly, 2003). Des études sur l’alimentation contrôlée ont démontré que les otaries, particulièrement les jeunes individus, qui consomment de grandes quantités de proies à faible teneur en gras telles que la goberge peuvent être incapables de maintenir leur masse corporelle (Rosen et Trites, 2000c; Azana, 2002). Dans la nature, ces jeunes individus seraient d’ordinaire toujours dépendants de leurs mères, qui pourraient alors avoir de la difficulté à assumer le coût énergétique élevé de la lactation (Winship et al.,2002; Pitcher et al.,2003). En conséquence, la disponibilité de proies de grande qualité près des roqueries semble être un facteur limitatif potentiel important.
On a démontré que la diversité du régime alimentaire était inversement corrélée avec la gravité des déclins subis par la population de l’Ouest, dans le golfe d’Alaska, les déclins les plus marqués étant survenus dans des secteurs où le régime alimentaire était le moins diversifié (Merrick et al.,1997). La diversité du régime alimentaire de la population de l’Est en croissance semble être élevée (Trites et al.,données non publiées). Les taux réduits de croissance corporelle (Calkins et al.,1998) et une corrélation directe entre l’état corporel et la proportion de femelles qui se rendent jusqu’aux derniers stades de la gestation (Pitcher et al.,1998) constituent une nouvelle preuve des contraintes alimentaires associées aux déclins abrupts qu’a connus la population de l’Ouest au cours des années 1980 (NMFS, 2007).
Les effets aigus qu’une réduction de la disponibilité des proies peut avoir sur les pinnipèdes sont manifestes à la lumière des déclins abrupts qui ont touché la production de petits chez les otaries de Californie et les otaries à fourrure du Nord sur l’île San Miguel, lesquels déclins ont coïncidé avec des événements El Niño (DeLong et Antonelis, 1991; Melin et DeLong, 1994; Melin et al.,1996; Melin et DeLong, 2000).
Les otaries de Steller consomment également nombre des mêmes proies que celles recherchées par d’autres prédateurs, y compris celles prélevées par l’homme (McAlister et Perez, 1976; Kajimura et Loughlin, 1988; Fritz et al.,1995; Wada, 1998; Trites et al.,1999). La pêche sélective pratiquée par l’homme peut également provoquer des changements dans les stocks de poissons (Pauly et al.,1998).
Prédation
La question de savoir si le forçage descendant découlant de la prédation exercée par les épaulards migrateurs peut également limiter les populations d’otaries a attiré beaucoup d’attention au cours des dernières années. En 1992, on a trouvé, dans l’estomac d’un épaulard échoué à Prince William, en Alaska, 14 étiquettes de nageoire qui avait été installées sur des petits de l’otarie de Steller. Même si aucune donnée sur les taux de prédation n’est disponible, des modèles ont révélé que les épaulards migrateurs pouvaient avoir un impact important sur les populations d’otaries de Steller et, en particulier, qu’ils pouvaient nuire au rétablissement des populations décimées (Barrett-Lennard et al.,1995).
Maniscalco et al. (2007) estiment que les épaulards consomment de 3 à 7 % de la population d’otaries de Steller dans les fjords Kenai chaque année et 11 % des petits nés au principal site étudié sur l’île Chiswell, ce qui peut être important dans le cas d’une espèce qui affiche un taux de productivité intrinsèquement faible. Les calculs préliminaires révèlent que, même si la prédation exercée par les épaulards représentait l’ensemble de la mortalité naturelle, la production annuelle nette des populations d’otaries de Steller en C.-B. et celle de l’ensemble de la population de l’Est ne pourraient, selon les approximations, assurer la survie que de 26 et de 77 épaulards respectivement (Olesiuk, données non publiées).
Plusieurs menaces peuvent affecter cette population en Colombie-Britannique. Par menaces, on entend toute activité (tant naturelle qu’anthropique) qui a causé, qui cause ou qui peut causer des dommages, des mortalités ou des changements comportementaux chez une espèce en péril ou, encore, la destruction, la dégradation ou la détérioration de son habitat à un point tel que des effets seront ressentis par la population.
Les effets des menaces sont souvent difficiles à différencier les uns des autres ou des facteurs limitatifs naturels. Ainsi, l’exposition aux contaminants peut rendre les animaux plus vulnérables aux maladies naturelles; les perturbations des otaries aux échoueries et aux roqueries peuvent inciter celles-ci à trouver refuge dans l’eau, où elles seront plus vulnérables aux attaques des épaulards. En outre, comme ces animaux se concentrent à un nombre limité de sites de reproduction, ils peuvent être plus vulnérables aux accidents catastrophiques ou aux menaces localisées affectant leur survie aux premiers stades de leur vie (p. ex. épuisement localisé des proies à cause de la pêche ou de perturbations).
La section 1.5.1 présente, sous la forme d’un tableau, un résumé de l’évaluation du risque exposant les menaces en fonction de leur impact sur les populations d’otaries de Steller. On a relevé l’« importance » et les « mesures d’atténuation potentielles » de chacune d’entre elles (tableau 1). Ces évaluations permettent d’établir l’ordre de priorité des mesures de gestion et autres recommandées pour éviter que cette population ne devienne menacée ou en voie de disparition et offrent également un aperçu de la faisabilité de l’atténuation d’une menace. Les définitions des termes utilisés pour qualifier l’ampleur des menaces sont présentées à l’annexe I (tableau 4).
La section 1.5.2 donne des descriptions détaillées de douze menaces historiques, actuelles et potentielles pesant sur la population d’otaries de Steller de la C.-B. ainsi que des incertitudes entourant les effets sur ces populations. Cette section contient les renseignements de base que l’on a utilisés pour établir l’importance globale concernant l’impact de chaque menace sur la population de l’Est (tableau 1).
Bien qu’il existe une incertitude considérable quant à l’impact global des menaces pesant sur l’otarie de Steller dans les eaux canadiennes, la croissance soutenue de la population d’otaries de Steller locale semble indiquer que ces menaces ne portent pas atteinte à la durabilité de ces populations. Cependant, avec un taux de croissance démographique d’environ 4,5 % par année, une augmentation relativement faible de la mortalité anthropique pourrait devenir un facteur important si les conditions dans lesquelles vit l’otarie de Steller se détérioraient ou si ce facteur était combiné à d’autres menaces.
On a évalué les menaces selon leur probabilité d’occurrence actuelle et la gravité de leur incidence sur la population en C.-B. En outre, on a incorporé la certitude d’occurrence d’un effet sur l’ensemble de la population de la C.-B. à l’évaluation afin de fournir une mesure du degré de confiance que l’on peut accorder aux classements de « l’importance » de la menace et de relever les menaces qu’il pourra être utile de surveiller ou d’étudier davantage afin d’éliminer des incertitudes ou de combler des lacunes dans les connaissances. Lorsque la certitude de l’occurrence d’un effet n’est pas démontrée, les données scientifiques relatives à d’autres pinnipèdes ou mammifères marins peuvent être considérées comme adéquates pour l’évaluation de l’importance d’une menace.
Le potentiel d’atténuation (tableau 1) renvoie à la probabilité qu’une mesure (future ou actuelle) atténuera ou empêchera de façon adéquate l’occurrence d’effets négatifs sur la population. Il convient de noter que le classement indiqué pour l’importance de la menace reflète les préoccupations actuelles relatives à des impacts découlant d’une menace présente et que les évaluations futures pourront entraîner des classements qui différeront de ceux dont il est question dans le présent document. En conséquence, l’importance d’une surveillance à long terme de la population ne peut être mise en doute.
Tableau 1. Résumé de la classification des menaces et du potentiel d’atténuation des menaces relevées pour l’otarie de Steller, population de l’est du Pacifique. Le potentiel d’atténuation renvoie à la probabilité qu’une mesure (future ou actuelle) puisse atténuer les effets négatifs sur la population ou empêcher leur occurrence. La présente évaluation se veut un reflet à jour de l’état des menaces pesant sur la population; à ce titre, le classement de l’importance de la menace peut changer au fil du temps. L’astérisque (*) signifie que la menace est naturellement présente dans la population (c.-à-d. qu’il s’agit d’un facteur limitatif dont les effets peuvent être amplifiés par l’activité humaine).
Menace |
Catégorie d’âge la plus vulnérable | Facteurs limitatifs qui sont susceptibles d’être affectés | Gravité de l’impact sur la population | Incertitude | Importance actuelle | Potentiel d’atténuation | |
| Juvéniles, femelles reproductrices |
Disponibilité des proies Impact direct : survie La limitation chronique des proies peut entraîner une baisse des taux de reproduction |
Potentiellement élevée | Moyenne | Modérée, potentiellement élevée |
Élevé | ||
| Juvéniles, femelles reproductrices |
Disponibilité des proies Possibilité de modification de la répartition Occurrence de maladies naturelles Impact direct : survie La limitation chronique des proies peut entraîner une baisse des taux de reproduction |
Potentiellement élevée | Moyenne | Modérée, potentiellement élevée |
Nul si la menace est attribuable à des fluctuations naturelles Faible si la menace est attribuable à des effets anthropiques sur le climat |
||
| Contaminants environnementaux | Polluants organiques persistants (POP) non réglementés, (p. ex. PBDE) |
Petits, femelles adultes |
Qualité des proies Vulnérabilité accrue aux maladies Réduction des taux de reproduction |
Modérée | Moyenne-élevée | Modérée | Faible-moyen |
| POP réglementés, (p. ex. DDT) | Petits, femelles adultes |
Qualité des proies Vulnérabilité accrue aux maladies Réduction des taux de reproduction |
Modérée | Moyenne-élevée | Faible-modérée | Faible-moyen | |
| Perturbations | Perturbations physiques lorsque les otaries sont dans un habitat terrestre | Petits aux roqueries | Survie des petits Comportements territoriaux et reproducteurs |
Faible aux échoueries Modérée aux roqueries |
Moyenne-élevée | Faible-modérée | Élevé |
| Perturbations acoustiques lorsque les otaries sont dans un habitat aquatique | Toutes | Utilisation de l’habitat (c.-à-d. éloignement des zones d’alimentation) Succès de l’alimentation La limitation chronique des proies peut entraîner une baisse des taux de reproduction |
Vraisemblablement faible |
Moyenne | Faible | Élevé | |
| Déversements de substances toxiques | Petits, femelles adultes aux roqueries pendant la saison de reproduction | Utilisation de l’habitat Impact direct : survie |
Faible Modérée aux roqueries de l’île Scott et du cap St. James |
Moyenne-élevée | Faible-modérée | Faible-moyen | |
| Emprisonnement accidentel – pêches et aquaculture | Inconnue | Impact direct : survie | Faible | Élevée | Faible | Moyen | |
| Emmêlement dans des débris marins | Juvéniles et subadultes | Réussite de l’alimentation Impact direct : survie |
Faible, effets potentiellement graves chez certains individus | Moyenne | Faible | Moyen | |
| Abattage illégal d’otaries | Juvéniles, adultes | Impact direct : survie | Inconnue | Élevée | Faible | Moyen | |
| Prédation par les épaulards* | Petits, juvéniles et adultes | Impact direct : survie | Potentiellement élevée | Moyenne | Faible | Nul | |
| Programmes d’abattage des prédateurs | A affecté autrefois toutes les classes d’âge Actuellement sans objet |
Impact direct : survie | Historiquement élevée Actuellement faible |
Faible | Historiquement élevée ActuellementNégligeable |
Élevé | |
| Prélèvements par les Premières nations | Toutes | Impact direct : survie | Faible | Élevée | Négligeable | Élevé | |
| Maladies et parasites* | Toutes | Les effets peuvent être amplifiés par les effets synergiques des menaces Impact direct : survie Réduction des taux de reproduction |
Inconnue | Moyenne-élevée | Inconnue | Faible | |
Diminution de la disponibilité des proies – concurrence exercée par les pêches
L’otarie de Steller consomme en grande partie les mêmes proies que celles recherchées par d’autres prédateurs, dont l’humain (McAlister et Perez, 1976; Kajimura et Loughlin, 1988; Fritz et al.,1995; Wada, 1998; Trites et al.,1999). Les prélèvements commerciaux peuvent décimer les populations locales et réduire la disponibilité des proies (Lowe et Fritz, 1997; Fritz et Brown, 2005), tandis que le prélèvement de la production excédentaire sur une base continue peut affecter la résilience et amplifier les effets des fluctuations naturelles des populations de proies. Comme les femelles qui allaitent doivent s’alimenter à proximité des roqueries pendant les premiers mois qui suivent la mise bas, la disponibilité des proies dans ces secteurs peut être essentielle à la survie des petits et de leurs mères.
Il est maintenant largement reconnu que le déclin qui a touché la population de l’Ouest a, au moins jusqu’à un certain point, été provoqué par un changement survenu dans le régime alimentaire, lequel changement a entraîné une réduction de la croissance corporelle, des taux de naissance et, en bout de ligne, de la survie (Calkins et Goodwin, 1988; Calkins et al.,1998; Pitcher et al.,1998; voir l’examen effectué par Trites et Donnelly, 2003). Cependant, le débat se poursuit concernant l’incidence relative des fluctuations naturelles des conditions environnementales, des changements dans les régimes climatiques et océaniques et des effets anthropiques qui peuvent découler du réchauffement planétaire, de la chasse à la baleine et des pêches commerciales (Pascual et Adkinson, 1994; Fritz et Ferrero, 1998; Pauly et al., 1998; Trites et al., 1999; Rosen et Trites, 2000a; Shima et al., 2000; Benson et Trites, 2002; Fritz et Hinkley, 2005; Trites et al., 2006; Trites et al., 2007). Pour de plus amples renseignements sur les proies, voir la section « Variabilité et changement au sein de l’environnement ».
Plusieurs pêches commerciales ciblent des espèces-proies que l’on sait importantes pendant l’été pour l’otarie de Steller en Colombie-Britannique : sardine, hareng, merluche, saumon et poissons de fond. Toutes ces espèces font l’objet d’une gestion quant aux taux de prise, lesquels taux sont considérés comme durables. On sait que l’otarie de Steller s’alimente d’une multitude d’espèces de proies, mais on ignore si des restrictions quant à la disponibilité d’une seule espèce de proies peuvent limiter la croissance de la population. La disponibilité générale des proies et la composition des espèces dans une région peuvent être modifiées par des facteurs naturels et anthropiques, c’est pourquoi les gestionnaires des pêches devront rechercher un équilibre entre la nécessité d’assurer la poursuite de la croissance de la population d’otaries de Steller et les besoins des pêcheurs. On connaît mal le régime alimentaire des otaries de Steller dans les saisons autres que l’été; lorsque d’autres renseignements seront disponibles, des espèces supplémentaires pourront être établies comme représentant des composants importants du régime alimentaire annuel des otaries de Steller en C.-B.
Le potentiel d’atténuation de cette menace est élevé (tableau 1) du fait que les prélèvements associés aux pêches peuvent être gérés directement par Pêches et Océans Canada. Cependant, comme l’otarie de Steller et les proies dont elle se nourrit sont des espèces transfrontalières, la prise de mesures d’atténuation adéquates peut nécessiter la collaboration supplémentaire des gestionnaires des pêches aux États-Unis.
À la lumière de la croissance exponentielle non limitée de la population d’otaries de Steller au cours des 45 dernières années, la concurrence pour les proies entre les pêches commerciales et cette population ne semble pas avoir eu d’effet sur cette dernière, et c’est pourquoi l’inquiétude est modérée en ce qui concerne les impacts. La réduction d’un approvisionnement en proies de grande qualité a eu des effets négatifs démontrés sur l’otarie de Steller – population de l’Ouest (Calkins et Goodwin, 1988; Calkins et al.,1998; Pitcher et al.,1998; voir l’examen effectué par Trites et Donnelly, 2003) et les besoins en matière de proies de l’otarie de Steller augmentent au fur et à mesure que la population de l’Est continue de s’accroître. Les impacts éventuels sur la population revêtent donc une certaine importance (tableau 1). Cette situation démontre l’importance de poursuivre la surveillance des pêches et des conditions océaniques et d’éclaircir les besoins saisonniers en matière de proies afin de prévoir toute augmentation de la concurrence entre l’espèce et les pêches à l’égard de la ressource; cet effort peut également contribuer à l’élaboration et à l’application de mesures de gestion appropriées.
Diminution de la disponibilité des proies – variabilité et changement au sein de l’environnemen
Le degré de changement survenant dans l’écosystème naturel (souvent par étapes discrètes que l’on appelle « changements de régime ») est fonction d’un certain nombre de facteurs, dont nombre sont mal compris, leur cause n’étant pas toujours apparente. Même si les changements de régime peuvent être d’origine anthropique, notamment le changement climatique mondial qui affecte les processus océanographiques (p. ex. changements dans la température des océans ou dans la répartition des espèces), des changements peuvent également survenir en raison de processus naturels tels que les oscillations décennales et les événements El Niño. La pêche peut également provoquer des changements dans les stocks de poissons et le fonctionnement des réseaux trophiques marins (Pauly et al.,1998).
Comme on l’a mentionné dans la section « Facteurs limitatifs », le changement climatique et les changements de régime à grande échelle peuvent affecter le biote dans l’ensemble du Pacifique Nord (Sinclair et al.,1994; Beamish et Bouillon, 1993; Sinclair et al.,1996; Anderson et al.,1997; Anderson et Piatt, 1999; Hare et al.,1999; McFarlane et al.,2000; Benson et Trites, 2002); or, de tels changements peuvent influer sur la répartition des proies de l’otarie de Steller en C.-B. et dans l’ensemble de l’aire de répartition de cette espèce (NMFS, 2007). L’augmentation des températures de l’océan découlant du changement climatique mondial pourrait entraîner un déplacement de l’aire de répartition de l’otarie de Steller vers le nord (NMFS, 2007) et, effectivement, l’espèce est en train de disparaître de l’extrémité sud de son aire de reproduction, tant sur la côte nord-américaine que sur la côte asiatique (Pitcher et al.,2007; Burkanov et Loughlin, 2007). Le centre de l’aire de répartition de la population reproductrice sur la côte ouest de l’Amérique du Nord s’est déplacé vers le nord, passant du fleuve Columbia (46,0 ºN) dans les années 1920 au centre de la C.-B. (51,5 ºN) en 2002 (Pitcher et al.,2007).
Les impacts aigus qu’une disponibilité moindre des proies peut avoir sur les pinnipèdes sont démontrés clairement par les déclins abrupts observés dans la production de petits chez les otaries de Californie et les otaries à fourrure du Nord sur l’île San Miguel, lesquels déclins ont coïncidé avec des événements El Niño (DeLong et Antonelis, 1991; Melin et DeLong, 1994; Melin et al.,1996; Melin et DeLong, 2000). L’apparition d’un changement de régime dans l’abondance des proies – les espèces-proies à forte teneur énergétique ayant été remplacées par des espèces à faible teneur énergétique (p. ex. le hareng remplacé par des gadidés) – est susceptible d’affecter les paramètres du cycle biologique des otaries (Trenberth, 1990; Springer, 1998; Benson et Trites, 2002; Trites et al.,2007) et de se traduire par un déclin des populations.
Actuellement, la population d’otaries de Steller de la C.-B. a dépassé les effectifs les plus élevés jamais atteints. Comme les populations continuent de s’accroître et que les besoins en matière de proies augmentent, l’otarie de Steller peut devenir plus vulnérable au manque de proies. L’importance des effets sur la viabilité de la population découlant d’un changement de régime ou du changement climatique est modérée. Cependant, si des changements extrêmes survenaient dans l’écosystème, entraînant une diminution de la disponibilité des proies, il y aurait alors lieu de s’inquiéter fortement de la possibilité d’impacts sur la population. En raison de la grande incertitude entourant l’occurrence/les effets d’un simple changement de régime ou les effets de modifications chroniques à long terme des conditions océaniques (imputables au changement climatique mondial), il est prudent d’assurer un suivi de la population. En établissant mieux quelles sont les proies importantes sur une base saisonnière pour l’espèce, nous pourrions mieux prévoir les impacts qu’aurait un changement de régime sur l’occurrence et la répartition des espèces-proies.
Contaminants environnementaux – Polluants organiques persistants
Les contaminants environnementaux persistants, comme les pesticides organochlorés (p. ex. DDT), les biphényles polychlorés (BPC), les dibenzodioxines polychlorées (PCDD), les dibenzofuranes polychlorés (PCDF) et les polybromodiphényléthers (PBDE) sont bioaccumulables dans les réseaux trophiques marins. L’accroissement des concentrations de ces substances chimiques au fur et à mesure que l’on gravit des échelons du réseau trophique prédispose de nombreux mammifères marins à une très forte contamination.
Dans certains cas, les concentrations de contaminants environnementaux persistants ont été associées à des effets négatifs sur la santé de pinnipèdes vivant en liberté. Des études menées sur le terrain laissent entrevoir l’existence de liens entre les concentrations de contaminants et des problèmes de reproduction (Helle, 1976a,b; Addison, 1989), des naissances prématurées (DeLong et al.,1973; Gilmartin et al.,1976; Martin et al.,1976), des anomalies congénitales (Arndt, 1973), des malformations squelettiques (Bergman et al.,1992), une immunosuppression (Mos et al., 2006) et la perturbation du comportement physiologique de la vitamine A et des hormones thyroïdiennes (Tabuchi et al., 2006; Mos et al., 2007). Des études d’alimentation menées sur des phoques communs vivant en captivité ont également démontré les effets néfastes des contaminants persistants sur les systèmes reproductifs, immunitaires et endocriniens (Brouwer et al.,1989; de Swart et al.,1994; Ross et al.,1995; Reijnders et al., 1986; Ross et al.,1996). Les résultats obtenus dans nombre de ces études correspondent au profil des effets observés chez des animaux de laboratoire exposés à des BPC ou à des dioxines (Ross et al., 1997; Ross, 2000).
Les petits allaités sont exposés à des concentrations particulièrement élevées de contaminants du fait que les produits chimiques solubles dans le gras, tels les BPC, leur sont facilement transférés par le lait riche en lipides de leur mère. (Hickie et al., 2005). Même si la plupart des contaminants persistants ne sont pas susceptibles d’afficher une toxicité aiguë, on considère qu’il s’agit de composés qui perturbent le système endocrinien (« perturbateurs endocriniens »; Colborn et al., 1993). À ce titre, ils peuvent altérer la croissance et le développement des animaux exposés. En outre, les effets peuvent demeurer inaperçus jusqu’à ce qu’un cofacteur de déclenchement n’entre en jeu, comme une période de jeûne ou une période de pénurie alimentaire qui peut accroître la mobilisation des substances chimiques liposolubles ou les contraintes imposées au système immunitaire (Jepson et al.,2005). Il pourrait, par exemple, s’agir d’un nouveau virus qui est introduit dans une population naïve de pinnipèdes chez laquelle les effets associés à la toxicité des contaminants peuvent entraîner une augmentation de la vulnérabilité, de la transmission de virus, de la gravité des maladies ou du taux de mortalité (Ross, 2002).
Comme cela a été démontré chez d’autres mammifères marins, les concentrations de contaminants chez les otaries de Steller (principalement des organochlorés) sont liées à l’âge et au sexe. Les concentrations les plus élevées s’accumulent chez les vieux mâles, tandis que les femelles transfèrent la majeure partie de leur charge en contaminants à leurs petits pendant la lactation (Lee et al.,1996). Barron et al. (2003) ont constaté que les concentrations de BPC chez les otaries de Steller semblaient être en déclin dans le golfe d’Alaska ces dernières années et que les concentrations actuelles ne constituent pas une menace; cependant, les concentrations de BPC observées dans les années 1980 étaient plus élevées chez les otaries de Steller que chez tout autre pinnipède et auraient pu poser un risque pour la santé.
En C.-B., on a effectué peu d’études systématiques des concentrations de contaminants chez les otaries de Steller, même si on a amorcé des études récemment pour évaluer les BPC et les PBDE. Les BPC, les dioxines, les furanes et les pesticides organochlorés sont considérés comme étant des contaminants anciens, réglementés depuis que des restrictions nationales et internationales ont été mises en œuvre concernant leur utilisation, leur production ou leur fabrication en tant que sous-produits. Cependant, de nombreux produits chimiques nouveaux et non réglementés sont considérés comme étant des sources de préoccupation « émergentes ». Parmi ces nouveaux produits, mentionnons les PBDE ignifuges, dont la concentration dans l’environnement double tous les quatre ans (Hites, 2004; Ross, 2006). Comme les PBDE ressemblent aux BPC, il est probable que l’on reconnaîtra de plus en plus que ces substances constituent une menace pour la santé de la faune, y compris celle de l’otarie de Steller.
Le poids de la preuve recueillie concernant les effets et la persistance des polluants réglementés et non réglementés sur les otaries de Steller soulève des inquiétudes quant à leurs effets sur la viabilité de la population. L’importance de cette menace est modérée1 du fait de la présence croissante des POP nouveaux et non réglementés dans l’environnement ainsi que leur usage global, tandis que l’importance de la menace posée par les contaminants anciens réglementés est considérée comme étant de faible à modérée (tableau 1). Cependant, on peut s’attendre à ce que la population soit plus vulnérable pendant les périodes caractérisées par une limitation du nombre de proies et par des contraintes nutritionnelles, durant lesquelles les lipides et leurs fortes concentrations de contaminants sont mobilisés. Comme des incertitudes subsistent quant à la charge toxique chez les otaries de Steller de la C.-B., la surveillance de cette menace et la prise de mesures d’atténuation visant les sources de pollution sont recommandées.
Même si les sources ponctuelles de contamination peuvent être réglementées et surveillées, la possibilité de mettre en œuvre des mesures d’atténuation concernant cette menace est considérée comme étant de faible à moyenne en raison des difficultés associées à la prise de mesures d’atténuation ou de gestion des sources de pollution diffuses et du transport aérien des contaminants sur de longues distances. En outre, les sources de contamination d’origine canadienne peuvent faire l’objet de mesures d’atténuation, tandis que les possibilités d’atténuation des sources de contamination situées en eaux internationales sont très faibles dans la perspective d’une gestion canadienne.
Perturbations sur l’habitat terrestre et à proximité de celui-ci
Une perturbation répétée des sites de reproduction ou des échoueries par des avions, des bateaux, des randonneurs, des ouvriers procédant à des travaux de construction ou des pêcheurs (p. ex. pêche en plongée à la panope ou à l’oursin) peut inciter les otaries à quitter temporairement les échoueries et les roqueries (Sandegren, 1970; Calkins et Curatolo, 1980; Johnson et al.,1989; Brown, 1997) et, éventuellement, à abandonner ces sites de façon permanente (Pike et Maxwell, 1958; Kenyon, 1962).
La perturbation des pinnipèdes aux échoueries par des navires est fonction d’une série de facteurs, y compris le type et le nombre de navires, leur vitesse et leur distance par rapport aux animaux (Henry et Hammill, 2001; Szaniszlo, 2005). Fait intéressant, il a été démontré que les embarcations à rames (telles que les kayaks) suscitent des réactions comportementales chez les pinnipèdes présents aux échoueries (Henry et Hammill, 2001), probablement en raison du risque de prédation évoqué par les rameurs (Deecke et al.,2002; Frid et Dill, 2002). Néanmoins, les otaries de Steller présentes aux sites d’alimentation hivernaux s’habituent souvent aux perturbations chroniques; ainsi, certaines échoueries sont situées dans des zones de fort trafic, à proximité de centres urbains majeurs tels que Vancouver et Victoria (Bigg, 1985; P. Olesiuk, comm. pers.). Les perturbations associées aux bruits aigus, comme les bruits de dynamitage ou de démolition, à proximité d’échoueries peuvent entraîner des réactions de panique chez les otaries de Steller et causer des perturbations localisées prononcées (par opposition aux perturbations touchant l’ensemble de la population).
Même si les animaux aux échoueries peuvent s’accoutumer à l’activité humaine, les otaries de Steller sont vulnérables aux perturbations touchant les roqueries. L’intrusion de l’homme dans les roqueries pendant la saison de reproduction (p. ex. pour recenser les animaux, capturer des petits afin de les marquer) semble perturber fortement les animaux et, souvent, incite ceux-ci à fuir dans l’eau ou vers des échoueries avoisinantes (Lewis, 1987; Scordino, 2006; Olesiuk, obs. non publiées). Les perturbations peuvent entraîner un accroissement de la mortalité des petits en raison de noyades, de piétinements ou d’une séparation d’avec leur mère. Le temps de retour aux échoueries après une perturbation causée, par exemple, par la présence de personnes procédant à une collecte d’excréments varie fortement, allant de quelques heures à deux semaines (Kucey, 2005; J. Etzkorn, Carmanah Lighthouse, C.-B., comm. pers.), tandis que l’effet d’une perturbation à une roquerie peut se faire sentir pendant un certain nombre d’années (Olesiuk, obs. non publiées). En outre, les perturbations peuvent avoir un coût énergétique pour les petits et les mères si les activités d’alimentation ou d’allaitement sont perturbées.
La fidélité aux sites de reproduction est un autre facteur associé aux perturbations survenant aux roqueries. Pendant les programmes d’abattage exécutés avant 1970, certains reproducteurs se sont vraisemblablement déplacés vers d’autres roqueries, mais la majorité des otaries ont continué à revenir aux sites qui avaient été fortement perturbés pendant de nombreuses années consécutives. À la roquerie du groupe Sea Otter, malgré l’abattage annuel intense qui s’y est déroulé, les otaries ont continué à utiliser la roquerie pendant 17 ans avant que la colonie soit complètement éradiquée. En conséquence, la biologie de la reproduction de l’otarie de Steller n’offre peut-être pas la souplesse nécessaire pour permettre à l’espèce de s’adapter aux perturbations survenant près des colonies reproductrices.
Comme on l’a mentionné précédemment, la gravité de l’impact sur la population est faible dans le cas des échoueries, mais est modérée dans le cas des perturbations survenant aux roqueries en raison de la fidélité au site, des coûts énergétiques potentiels pour les mères et les petits ainsi que de la survie des petits (tableau 1). Le potentiel d’atténuation de cette menace est élevé. À l’heure actuelle, l’accès aux roqueries fait l’objet d’une surveillance stricte, et il faut disposer de permis pour entrer sur une roquerie reconnue. La perturbation des pinnipèdes par les navires est gérée grâce à des lignes directrices s’appliquant à l’observation des mammifères marins. D’après les critères énoncés ci-devant, l’importance de cette menace est considérée comme étant de faible à modérée. Il convient de noter que la réglementation américaine interdit l’accès dans une zone s’étendant sur trois milles autour des roqueries (dans l’ouest de l’Alaska), ce qui illustre bien la nature délicate de la perturbation aux roqueries.
Perturbations acoustiques dans l’habitat aquatique
Les activités associées à la prospection pétrolière et gazière, à l’exploitation de sources d’énergie de rechange (p. ex. énergies éolienne et marémotrice) ainsi que d’autres activités d’extraction des ressources (p. ex. méthanol) peuvent perturber les animaux du fait qu’elles produisent des bruits sous-marins chroniques (découlant des activité de navigation ou de construction) et aigus (découlant des relevés sismiques).
Les perturbations acoustiques, comme les explosions et les bruits de sonars sismiques ou tactiques militaires, peuvent inciter les animaux à quitter leurs aires d’alimentation et perturber les comportements d’alimentation. Les normes canadiennes sur les relevés sismiques contribueront à l’atténuation de cette menace. L’Énoncé des pratiques canadiennes d’atténuation des ondes sismiques en milieu marin précise des normes minimales qui doivent être respectées pendant la réalisation de relevés sismiques en mer dans toutes les eaux marines libres de glace du Canada (http://www.dfo-mpo.gc.ca/oceans-habitat/oceans/im-gi/seismic-sismique/statement-enonce_f.asp). Comme les otaries de Steller sont capables de faire surface et de quitter l’eau pour éviter les contraintes dues aux bruits aigus, l’importance des perturbations par les bruits aigus aux sites d’alimentation est faible. Cependant, on observe actuellement un intérêt pour l’expansion des activités de prospection et d’extraction des combustibles fossiles au large; par conséquent, l’évaluation de cette menace devra être effectuée de façon continue lorsque de nouvelles données sur l’occurrence et la fréquence de ces activités et sur leurs effets sur les roqueries et les aires d’alimentation des otaries de Steller seront disponibles. Il faudra poursuivre l’examen des projets de développement et de prospection afin de s’assurer que les besoins en matière d’habitat de l’otarie de Steller seront pris en considération dans les plans de développement durable.
Les contraintes associées aux bruits chroniques dans les aires d’alimentation importantes et à proximité des roqueries peuvent avoir un effet à long terme sur les taux de survie des otaries de Steller et leur condition corporelle; cependant, étant donné l’éloignement des roqueries actuelles, l’inquiétude vis-à-vis de cette menace demeure faible (tableau 1). La prise en considération des roqueries au moment de l’aménagement d’installations industrielles devrait contribuer à atténuer cette menace chronique potentielle. L’augmentation générale du trafic maritime (bateaux motorisés et à rames) le long de la côte de la C.-B. a accru le nombre et la fréquence des visites aux échoueries. La perturbation acoustique dans l’eau (ou visuelle) causée par les activités de navigation pourrait contribuer à accroître les coûts énergétiques si les activités d’alimentation devaient être perturbées à long terme. La possibilité d’atténuer la perturbation acoustique sous-marine, en particulier aux environ des roqueries, est élevée du fait que deux des roqueries sont situées à l’intérieur de réserves de parc national et du fait que les lignes directrices et protocoles concernant la perturbation des mammifères marins par les bruits aigus pourraient faire l’objet d’un examen et d’une révision.
Déversements de substances toxiques
Les otaries peuvent être affectées par des accidents catastrophiques tels que des déversements de substances toxiques (St. Aubin, 1990), bien que l’impact de tels événements sur l’ensemble d’une population ait rarement été établi. La principale menace est vraisemblablement associée au contact avec des accumulations de pétrole lourd lorsque la source du déversement se trouve à proximité d’habitats importants tels que des roqueries et des échoueries et, à un degré moindre, par absorption cutanée, ingestion accidentelle de pétrole par voie directe ou par la consommation de proies, exposition à des vapeurs et salissure partielle du pelage par du pétrole frais (Smith et Geraci, 1975; Engelhardt et al.,1977; Engelhardt, 1987; St. Aubin, 1990). Comme les otaries possèdent une couche de gras sous-cutanée isolante, la présence de pétrole sur leur fourrure n’a aucune incidence sur la thermorégulation de leur organisme. Des otaries de Steller présentant du goudron dans leur gorge et autour de leurs lèvres, de leur mâchoire et de leur cou ont été observées en Alaska (Calkins et Pitcher, 1982).
Lors du déversement de pétrole de l’Exxon Valdez survenu dans le détroit de Prince William en 1989, le pétrole n’a pas persisté sur les fourrures des otaries de Steller aussi longtemps qu’il n’est demeuré sur celui des phoques communs (Calkins et al.,1994a), mais on a observé des otaries à proximité du pétrole déversé, et la découverte de métabolites dans leur sang a confirmé qu’elles avaient été exposées aux hydrocarbures. Les naissances prématurées ont été plus courantes et la production de petits a été quelque peu inférieure dans l’année qui a suivi le déversement; toutefois, en raison des données limitées recueillies au cours de la période précédant cet accident et du déclin continu de la population qui avait déjà lieu dans le secteur, il a été difficile d’évaluer l’importance statistique de l’impact de ce déversement (Calkins et al.,1994b; Loughlin et al.,1996).
On a observé plusieurs otaries de Steller avec de petites zones de fourrure souillées par le pétrole pendant le déversement du Nestucca qui s’est dispersé le long de la côte ouest de l’île de Vancouver en 1988 (Harding et Englar, 1989). Comme la population est largement répartie le long de l’ensemble de la côte de la C.-B., la menace potentielle associée aux déversements accidentels est localisée et n’aura pas d’impact sur la population entière. Cependant, si un déversement devait toucher une roquerie pendant la saison de reproduction, un impact important sur la population pourrait survenir. Si l’on considère que plus de 70 % de la production de petits en C.-B. a lieu sur les îles Scott, un déversement de pétrole dans ce secteur pendant la saison de reproduction pourrait avoir un impact important sur les animaux reproducteurs.
Comme on considère qu’un impact touchant l’ensemble de la population est improbable, l’inquiétude vis-à-vis des déversements catastrophiques affectant la population est considérée comme faible (tableau 1). Cependant, étant donné qu’un déversement survenant près d’une roquerie (p. ex. les îles Scott) pendant la saison de reproduction pourrait avoir un impact sur une forte proportion des animaux présents sur le site, un degré d’importance modéré est ajouté pour les impacts survenant aux roqueries pendant la saison de reproduction (tableau 1). Comme les déversements sont des événements accidentels, le moment et l’emplacement où ceux-ci ont lieu sont difficiles à prévoir. Des règlements et des mesures sont actuellement en vigueur au Canada pour limiter le risque de déversements accidentels (p. ex. Loi sur le transport des marchandises dangereuses) et pour atténuer les effets de ceux-ci (restauration de l’habitat et autres mesures). Le potentiel d’atténuation de cette menace est considéré comme étant de faible à moyen étant donné la difficulté inhérente à l’application des mesures de nettoyage à la suite d’un déversement et le faible degré de réussite obtenu dans ce domaine (Graham, 2004), particulièrement dans les zones isolées et éloignées.
Emprisonnement dans les engins de pêche ou les installations aquicoles
On observe des mortalités accidentelles d’otaries de Steller dans diverses pêches (en particulier les pêches au filet maillant dérivant visant le saumon), mais la couverture des pêches par les observateurs n’est pas suffisamment complète pour nous permettre d’assurer un suivi adéquat des prises accessoires. Des otaries peuvent être emprisonnées dans des chaluts ou capturées (piégées) dans des filets maillants et des filets maillants dérivants, où elles périront noyées (Loughlin et Nelson, 1986). Malheureusement, lorsqu’une otarie est emprisonnée, la possibilité de la récupérer pour la retourner dans son milieu est extrêmement faible, tant sur le plan technique que sur le plan pratique. Les prises accessoires annuelles dans les eaux américaines au cours des dernières années ont été estimées à environ 25 individus par année (Loughlin et York, 2000; Angliss et Outlaw, 2007). Cependant, aucune estimation similaire n’est disponible pour les pêches pratiquées dans les eaux de la C.-B.
Les taux d’emprisonnement dans les installations piscicoles sont actuellement établis par l’entremise de mécanismes de déclaration volontaire dans la région du Pacifique, ce qui peut limiter l’information concernant ces types d’interaction. D’après les déclarations volontaires compilées depuis 2004, un seul individu a été identifié comme étant une otarie de Steller (noyée après s’être emprisonnée) et 12 autres pinnipèdes ont été emprisonnés, sans toutefois pouvoir être identifiés2.
La précision des déclarations des espèces capturées n’a pas été établie; cependant, une formation menée par l’industrie est en cours. La mise en œuvre de mécanismes normalisés de déclaration volontaire vise à améliorer la déclaration et la consignation de ces occurrences. On obtiendra ainsi de meilleurs renseignements sur les taux d’emprisonnement des otaries de Steller dans les installations aquicoles. À l’heure actuelle, on ne recense aucune installation aquicole en Alaska, et c’est pourquoi aucune donnée n’est disponible à des fins de comparaison avec des cas d’emprisonnement d’otaries de Steller ailleurs dans le monde.
L’amélioration des efforts des producteurs aquicoles pour signaler les cas d’emprisonnement de mammifères marins a entraîné une augmentation des déclarations; cependant, cette situation n’indique pas nécessairement une augmentation générale des nombres de cas. Selon les estimations actuelles de la population d’otaries de Steller en C.-B., l’importance de cette menace est considérée comme faible en C.-B. (tableau 1). Cependant, des mesures préventives, y compris l’établissement des installations aquicoles à une certaine distance des échoueries et l’apport de modifications aux engins de pêche et aux installations d’élevage, ainsi que des mesures non létales pour éviter les accidents aux installations aquicoles sont mises en œuvre afin de limiter les risques d’emprisonnement. Il faut procéder à un examen et une révision continuels de ces méthodes afin de limiter les interactions létales.
Emprisonnement dans des débris marins
La présence grandissante de débris synthétiques (fragments de filet, sacs de plastique et bandes d’emballage, etc.) constitue un problème de plus en plus important à l’échelle mondiale et est responsable en partie des déclins observés chez d’autres espèces de pinnipèdes (Fowler et Merrell, 1986; Fowler, 1988). Les débris tels que les fragments de filet et les bandes d’emballage peuvent demeurer pris autour du cou des otaries de mer, ce qui provoque de l’abrasion et de profondes coupures dans les tissus des otaries lorsqu’elles grandissent. Parfois, des otaries de Steller consomment des poissons pris à des engins de pêche à la traîne, et il n’est pas rare de voir des otaries qui ont avalé des hameçons qui se sont accrochés à l’intérieur de leur tube digestif, avec un flasheur sortant de leur bouche. Malheureusement, il n’est pas possible ni sur le plan technique ni sur le plan pratique de récupérer les otaries emmêlées dans des débris marins pour les retourner dans leur milieu.
Les taux d’emprisonnement pour les otaries de Steller en Alaska ont été estimés à environ 0,07 % chez les adultes, les bandes d’emballage et les débris de filet étant les articles les plus fréquents (Calkins, 1985; Mate, 1985; Loughlin et al.,1986; Stewart et Yochem, 1987; Fowler, 1988). Au cours de campagnes de recherche menées récemment dans les eaux de la C.-B., on a dénombré qu’environ 0,2 % des individus avaient des débris pris autour de leur cou ou des engins de pêche accrochés à leur bouche ou dans leur gorge (Olesiuk, donnée non publiées). Cependant, comme l’a fait remarquer Fowler (1988), la majeure partie des débris présents en mer ou échoués sur le rivage sont trop gros pour que les animaux puissent les transporter; ainsi, le nombre d’individus observés avec des débris pourrait ne représenter qu’une petite fraction du nombre réel d’animaux emprisonnés et noyés en mer. On n’a observé aucun petit ni aucun jeune de l’année emprisonné dans des débris, et on ne sait pas précisément si ces groupes d’âge sont capables d’éviter les débris ou si l’emprisonnement d’animaux de plus petite taille se traduit par une mort certaine. La majeure partie des débris marins documentés comme cause d’emprisonnement sont associés à des engins de pêche perdus. L’apport de modifications aux engins pourrait réduire le risque d’emprisonnement ayant des conséquences négatives et être considéré comme une mesure d’atténuation potentielle relativement à cette menace.
La gravité de l’impact des emprisonnements dans des débris marins sur l’ensemble de la population d’otaries de Steller demeure inconnue; cependant, comme des cas d’emprisonnement sont observés, il est clair que cette menace affecte une certaine partie de la population. Il faut effectuer d’autres recherches pour combler les lacunes dans les connaissances concernant le taux d’emprisonnement et l’effet de ces débris à l’échelle de la population. Actuellement, l’importance de cette menace est faible (tableau 1).
Abattage illégal
L’abattage illégal et non documenté d’otaries de Steller a vraisemblablement lieu en C.-B., en particulier depuis que l’espèce est perçue comme ayant un impact négatif sur les stocks de poissons et depuis que l’on sait qu’elle s’attaque aux poissons élevés dans les installations piscicoles. Plusieurs cas d’abattage illégal ont été documentés (MPO, données non publiées), et des individus peuvent également être abattus en violation des paramètres réglementaires que doivent respecter les détenteurs de permis (p. ex. abattage des prédateurs ou prélèvements de subsistance). Cependant, on ne dispose actuellement pas de données sur ces activités, et les incidents qui se produisent dans des lieux éloignés sont évidemment plus difficiles à documenter ou à surveiller. Dans certaines zones éloignées, des activités de pêche ont lieu près de roqueries et d’échoueries, ce qui amplifie les interactions entre les pêches et les otaries du fait que ces dernières s’habituent à se rendre aux sites de rejet des abats et d’autres restes de poissons pour s’en nourrir.
On connaît mal l’ampleur de l’abattage illégal des pinnipèdes, et l’impact qu’a une telle activité sur les populations demeure inconnu. Cependant, à la lumière des récentes estimations de l’abondance de l’otarie de Steller en C.-B. (MPO, 2008d), il est peu probable que cette menace ait actuellement un impact sur la viabilité de la population (tableau 1). L’atténuation de cette menace passe par la tenue d’activités de vulgarisation et de communication avec les parties touchées, par des activités d’éducation pour modifier certaines pratiques (p. ex. rejets d’abats, accoutumance des animaux aux engins de pêche) ainsi que par une surveillance et une application accrues de la réglementation à proximité des échoueries et des roqueries utilisées par les otaries de Steller. À ce titre, le potentiel d’atténuation de cette menace est modéré.
Prédation par les épaulards
Comme on l’a vu dans la section « Facteurs limitatifs », les épaulards sont des prédateurs importants qui peuvent limiter les populations d’otaries de Steller. Les taux de prédation des otaries de Steller peuvent être accrus en raison des effets synergiques d’autres menaces ou facteurs limitatifs. La possibilité que les taux de prédation par les épaulards soient modifiés par la variabilité environnementale, des changements dans la disponibilité des proies (c.-à-d. l’augmentation de la distance à franchir pour s’alimenter) ou une incidence accrue des maladies peut accroître l’impact de cette menace naturelle sur la viabilité de la population d’otaries de Steller en C.-B. Des impacts sur d’autres proies des épaulards, comme les phoques communs, peuvent se traduire par un changement de proies chez les épaulards et un accroissement de la prédation visant les otaries. Les perturbations qui forcent les animaux à fuir dans l’eau ou à gagner d’autres sites peuvent également accroître leur exposition aux épaulards. En outre, la tendance à la hausse observée dans la croissance de la population des épaulards migrateurs (Ford et al.,2007) pourrait entraîner une augmentation du taux de prédation. Des modèles indiquent que les épaulards migrateurs pourraient avoir un impact important sur les populations d’otaries de Steller et, en particulier, qu’ils pourraient empêcher le rétablissement de populations décimées (Barrett-Lennard et al.,1995). Les calculs préliminaires révèlent que même si la prédation exercée par les épaulards était à l’origine de l’ensemble de la mortalité naturelle chez l’otarie de Steller, la production annuelle nette de la population d’otaries de Steller en C.-B. ainsi que de l’ensemble de la population de l’Est ne pourraient soutenir en gros que 26 et 77 épaulards respectivement (P. Olesiuk, données non publiées).
Comme la population d’otaries de Steller connaît actuellement une croissance non limitée et que d’autres proies, comme le phoque commun, présentent également des effectifs élevés, l’importance de la menace que représente un déclin démographique provoqué par la prédation est minimale à l’heure actuelle (tableau 1). Comme il est extrêmement improbable que des mesures d’atténuation des interactions prédateurs-proies soient mises en œuvre, la surveillance des populations d’otaries de Steller et d’épaulards migrateurs nous aidera à déterminer les tendances à long terme de l’abondance et la répartition de ces deux espèces dans les eaux de la C.-B. et dans l’ensemble de leurs aires de répartition.
Programmes d’abattage des prédateurs
Pendant la majeure partie du XXe siècle, le principal facteur limitatif des populations d’otaries de Steller le long de la côte ouest de l’Amérique du Nord a été la mise en œuvre de programmes d’abattage des prédateurs. En C.-B., les programmes gouvernementaux d’abattage des prédateurs ont entraîné la disparition d’une roquerie importante dans le groupe Sea Otter à la fin des années 1930, et le nombre d’otaries se reproduisant aux autres roqueries a été réduit à environ le quart des effectifs historiques à la fin des années 1960 (Bigg, 1985). Dans l’État de Washington, le gouvernement ayant offert une prime pour l’abattage des otaries de Steller, l’abondance de cette espèce sur la côte de cet État est passée de plusieurs milliers d’individus au début des années 1900 à un peu moins d’une centaine d’individus à la fin des années 1940. D’importantes campagnes d’abattage avec primes ont également eu lieu en Oregon dans les années 1920 (Pearson et Verts, 1970), et la perturbation de même que l’abattage dont les chasseurs et les pêcheurs étaient les auteurs ont également réduit l’abondance de l’otarie de Steller et, selon toute apparence, entraîné la disparition de plusieurs sites de reproduction en Californie (Rowly, 1929). En 1970 au Canada et en 1972 aux États-Unis, la population d’otaries de Steller de l’Est était déjà fortement décimée lorsque l’espèce a été protégée en vertu de la Loi sur les pêches et la Marine Mammal Protection Act respectivement. La seule partie de l’aire de répartition de la population de l’Est qui a échappé aux vastes campagnes d’abattage est le sud-est de l’Alaska, aucun registre ne démontrant que l’espèce se reproduisait ou était abondante dans cette région au début des années 1900.
De 1990 à 2003, le programme d’abattage réglementaire des prédateurs aux installations piscicoles de la C.-B. a représenté l’une des principales sources connues de mortalité anthropique chez l’otarie de Steller dans le Pacifique Nord (Angliss et al.,2001; Jamieson et Olesiuk, 2001). Les rapports trimestriels déposés par les détenteurs de permis indiquent qu’en tout 362 otaries de Steller et 21 otaries d’espèce inconnue ont été abattues de 1990 (lorsque les premiers permis ont été délivrés) à 2003. Le nombre d’otaries de Steller abattues chaque année a été faible au début (moins de 10 en moyenne), mais a grimpé à la fin des années 1990 et a culminé à 91 en 1999 (Jamieson et Olesiuk, 2001), vraisemblablement en raison d’un changement dans la répartition hivernale des otaries de Barkley vers le bras Clayoquot (P. Olesiuk, comm. pers.). En 2004, on a modifié les conditions rattachées aux permis afin d’épargner les otaries de Steller et, ainsi, éliminer l’utilisation de moyens létaux contre cette espèce du fait que cette dernière avait été désignée comme étant préoccupante par le COSEPAC. Cependant, nombre des quelque 100 installations salmonicoles exploitées dans les eaux de la C.-B. possèdent des permis les autorisant à utiliser des moyens létaux pour éliminer les phoques communs et les otaries de Californie, et il est possible que des otaries de Steller aient été abattues en raison d’une erreur d’identification de l’espèce. Étant donné le faible nombre d’individus actuellement abattus, l’importance de cette menace est considérée comme négligeable (tableau 1).
Prélèvements par les Premières nations
Les peuples autochtones de la C.-B. chassent depuis toujours l’otarie de Steller à des fins alimentaires (Bigg, 1985) et continuent d’utiliser leurs moustaches dans certains costumes cérémoniels traditionnels. L’utilisation des otaries par les Premières nations semble avoir décliné pendant les années 1800, et la viande d’otarie ne représente pas une partie importante de leur régime alimentaire depuis le début des années 1900 (Bigg, 1985). On ignore si la chasse de subsistance pratiquée avant les années 1900 a contribué à régulariser la population d’otaries de Steller en C.-B.
La chasse autochtone de l’otarie de Steller à des fins de subsistance et culturelles a cours de façon occasionnelle au Canada, mais les taux de prélèvement sont inconnus. Les Premières nations peuvent chasser les otaries sans permis; néanmoins, Pêches et Océans Canada travaille avec les Premières nations afin de les inciter à utiliser des permis communautaires assortis de limites sur les prélèvements afin de s’assurer que ces derniers ne dépassent pas les seuils de la durabilité. Étant donné qu’aucun permis de chasse commercial n’est délivré pour l’otarie de Steller en C.-B. et que les prélèvements de subsistance sont très limités, l’importance de cette menace est considérée comme étant négligeable (tableau 1). Le maintien des communications avec les groupes des Premières nations intéressés par la chasse aux pinnipèdes nous aidera à évaluer l’importance future des impacts que les prélèvements pourraient avoir sur l’ensemble de la population.
Maladies et parasites
Les tests de dépistage menés sur les otaries de Steller de la C.-B. et de l’Alaska ont donné des résultats positifs pour plusieurs agents pathogènes (Calkins et Goodwin, 1988; Sheffield et Zarnke, 1997; Burek et al.,2003, 2005; Lambourn et al.,2006) et, en général, les parasites sont courants chez les individus de l’espèce (Dailey et Hill, 1970; Dailey et Brownell, 1972; Fay et Furman, 1982; Shults, 1986; Gerber et al.,1993; cités dans NMFS, 2007). Même si les parasites et les maladies peuvent avoir peu d’impacts sur des animaux en santé, leurs effets peuvent être importants s’ils sont combinés avec d’autres facteurs d’agression (Haebler et Moeller, 1993). Les facteurs d’agression anthropiques peuvent également accroître l’incidence des maladies ou provoquer l’introduction d’agents pathogènes étrangers dans la population.
Les agents pathogènes et les maladies provenant de sources terrestres ou d’espèces exotiques constituent une préoccupation en ce sens qu’ils peuvent exposer la population à de nouveaux contaminants biologiques. Les rejets d’eaux d’égouts, les eaux de pluie et les eaux de ruissellement provenant des terres agricoles peuvent jouer un rôle important comme vecteurs de pathogènes ou de maladies, comme c’est le cas des programmes de réintroduction-réhabilitation plurispécifiques qui englobent des pinnipèdes.
La limitation des sources terrestres d’agents pathogènes et de l’exposition à des espèces exotiques affectant les mammifères marins contribuera à réduire le risque de transmission de maladies exotiques aux otaries de Steller de la C.-B. La prise de mesures pour limiter le risque d’exposition des pinnipèdes réhabilités à des agents pathogènes portés par des mammifères terrestres peut réduire la probabilité de transmission de maladies aux populations de pinnipèdes. Qui plus est, la gestion des sources de rejets et des eaux de ruissellement peut contribuer à atténuer cette menace. Cependant, étant donné la forte incertitude entourant les effets des maladies, la principale mesure à prendre est de combler les lacunes dans les connaissances concernant les sources terrestres d’agents pathogènes et leurs effets potentiels sur les otaries de Steller.
Les effets des menaces et des facteurs limitatifs peuvent difficilement être distingués les uns des autres, ce qui rend les conclusions concernant les causes des déclins des populations souvent difficiles à confirmer. Bien qu’il existe une incertitude considérable quant à l’impact total des menaces pesant sur les otaries de Steller dans les eaux canadiennes, la croissance continue de la population locale d’otaries de Steller semble indiquer que celle-ci se situe actuellement à l’intérieur des limites de la durabilité et que les effets individuels ou combinés des menaces et des facteurs limitatifs ne sont pas suffisamment importants pour provoquer un déclin de la population ou limiter la croissance de celle-ci. Néanmoins, avec un taux de croissance de la population inférieur à 5 % par année, une augmentation relativement faible de la mortalité anthropique pourrait devenir un facteur important si les conditions dans lesquelles les otaries de Steller vivent se détérioraient ou si cette mortalité était combinée avec d’autres menaces. En conséquence, on ne peut insister suffisamment sur l’importance d’exécuter des programmes de recherche ciblés visant à combler les lacunes dans les connaissances et sur la surveillance à long terme de la population et des menaces relevées.
Mesures de gestion des prélèvements
La conservation et la gestion des otaries de Steller au Canada relèvent des dispositions de la Loi sur les pêches (1985) et, de façon plus précise, du Règlement sur les mammifères marins (1993) promulgué en vertu de cette Loi. À l’exception des Premières nations, personne ne peut chasser ou perturber un mammifère marin, à moins d’avoir reçu l’autorisation explicite de le faire en vertu d’un permis. Selon l’article 6, les Premières nations peuvent chasser les otaries sans permis à des fins alimentaires, sociales ou cérémoniales; cependant, elles utilisent couramment des permis communautaires assortis de limites sur les prélèvements afin de s’assurer que ces derniers ne dépassent pas les seuils de la durabilité.
Depuis 1970, l’approche de gestion appliquée aux pinnipèdes en Colombie-Britannique n’autorise pas les prélèvements commerciaux ou l’abattage de ces animaux. Les seuls permis délivrés pour l’abattage d’otaries visent à protéger les stocks d’exploitations aquicoles et les bassins à harengs de la prédation. Des prélèvements limités sont permis pour les « animaux nuisibles » tels que les définit le Règlement sur les mammifères marins (un permis est exigé selon l’alinéa 26.1 (1) c); de plus, on surveille les prélèvements pour s’assurer qu’ils respectent les seuils de la durabilité. À la suite de la désignation de l’otarie de Steller en tant qu’espèce préoccupante par le COSEPAC en 2003, les permis d’abattage des prédateurs (phoques nuisibles) délivrés depuis 2004 en vertu de l’alinéa 26.1 (1) c) interdisent l’abattage des otaries de Steller.
Protection contre les perturbations
L’article 7 du Règlement sur les mammifères marins (Loi sur les pêches) interdit toute perturbation des mammifères marins à moins d’une autorisation en vertu d’un permis de pêche ou d’un permis scientifique. Les modifications proposées à ce règlement permettront aussi l’interdiction précise de certaines formes de perturbations telles que le fait de nager avec des mammifères marins ou de les nourrir. En outre, on a établi des lignes directrices (« Respectez les baleines! Directives pour l’observation de la faune aquatique à l’intention des plaisanciers et des observateurs », Paddlers et Viewers, 2006) pour corriger les problèmes de perturbations associées aux approches sur terre ou en mer à faibles distances. Des programmes de gestion et d’éducation, dont le but est d’assurer la conformité aux lignes directrices, ont été mis en œuvre pour l’industrie de l’écotourisme (p. ex. Whale Watch Operators Association Northwest Best Management Practices [ www.nwwhalewatchers.org ]) et le public.
Protection de l’habitat
La Loi sur les pêches (art. 35 et 36) contient des dispositions pour assurer la protection de l’habitat et prévenir la pollution ou la dégradation de l’habitat des mammifères marins. Les colonies reproductrices d’otaries de Steller aux îles Triangle et Beresford sont situées dans les réserves écologiques Anne Vallee (île Triangle) et de l’île Beresford respectivement. Ces réserves ont été créées en 1971 par le gouvernement de la Colombie-Britannique afin de protéger la biodiversité. Les réserves écologiques ne sont accessibles qu’aux personnes qui disposent d’un permis. Environnement Canada (Service canadien de la faune) dirige une initiative du gouvernement fédéral visant à créer une aire de faune marine (AFN) aux îles Scott, ce qui permettra l’atteinte des objectifs de la Loi sur les espèces en péril en assurant la protection de l’habitat de plusieurs espèces en péril inscrites sur la liste fédérale, dont l’otarie de Steller.
La colonie reproductrice d’otaries de Steller de cap St. James est située dans la réserve de parc national et le site du patrimoine haïda Gwaii Haanas, lesquels sont administrés dans le cadre d’une coopération entre l’Agence Parcs Canada et le Conseil de la nation haïda. On propose, en vertu de la Loi sur les parcs nationaux du Canada et de la Loi sur les aires marines nationales de conservation du Canada, d’agrandirl’aire marine nationale de conservation (AMNC) Gwaii Haanas, située au sud des îles de la Reine-Charlotte, de 10 km vers le large à partir de la réserve de parc national et du patrimoine haïda Gwaii Haanas pour protéger l’habitat marin de l’otarie de Steller dans cette région. Les AMNC sont gérées en fonction d’une utilisation durable et sont protégées des activités industrielles telles que l’immersion de déchets en mer, l’exploitation minière ainsi que l’exploration et l’exploitation des ressources gazières et pétrolières.
De nombreuses échoueries d’otaries de Steller de la C.-B. sont également protégées par des parcs nationaux ou provinciaux, comme les réserves de parc national Pacific Rim et des Îles-Gulf et la réserve écologique Race Rocks.
On examine actuellement l’Ordre du Commandement maritime pour la prise de mesures d’atténuation à l’intention des mammifères marins du ministère de la Défense nationale (MDN, 2007) afin de limiter les perturbations associées à l’utilisation des sonars tactiques. Par ailleurs, le Parlement a récemment adopté le Règlement sur les polybromodiphényléthers (juillet 2008). Cet outil réglementaire limite la fabrication et l’utilisation de divers types de PBDE au Canada et se veut un premier pas vers la réduction à long terme des effets toxiques de ces substances dans l’environnement. En outre, le Règlement sur la prévention de la pollution par les navires et sur les produits chimiques dangereux de la Loi sur la marine marchande du Canada vise la pollution et les débris en milieu marin. Ces codes de pratique et outils réglementaires peuvent contribuer à atténuer les effets sur l’habitat de l’otarie de Steller.
Après l’entrée en vigueur de la LEP en 2003, plusieurs programmes de rétablissement et plans de gestion de mammifères marins « en péril » ont été élaborés. Ces documents recommandent des mesures pour le rétablissement, la protection et la gestion des espèces de mammifères marins inscrites. Dans un contexte plus vaste, ces mesures de gestion peuvent également profiter aux otaries de Steller de la population de l’est du Pacifique vivant en C.-B. Se reporter à la section 4.0, « Plans connexes », pour connaître les programmes de rétablissement et les plans de gestion pertinents ainsi que les mesures associées à la protection et à la gestion des otaries de Steller en C.-B.
La Direction générale de la conservation et de la protection de Pêches et Océans Canada prend des mesures et, au besoin, mène des enquêtes lorsqu’on signale des prélèvements létaux non autorisés (y compris des tentatives à cet égard) et des perturbations de mammifères marins dans le cadre d’activités générales. La mise en application ciblée de la réglementation en matière de perturbation est axée sur l’observation des baleines, mais touche également la perturbation des pinnipèdes, en particulier lorsqu’ils sont sur une échouerie située à proximité de secteurs habités par l’homme. Cependant, les endroits où ont lieu ces activités ne chevauchent pas de manière importante l’aire de répartition de l’otarie de Steller.
L’information servant à soutenir l’application de la réglementation et la prise de mesures en cas d’incidents est recueillie par le Marine Mammal Response Network (MMRN) de la C.-B. (1-800-465-4336). Ces initiatives permettent la collecte de données sur les incidents mettant en cause des pinnipèdes et permettent la prise de mesures d’intervention ponctuelles.
Étant donné la répartition transfrontalière et la grande mobilité des otaries de Steller, les évaluations des populations doivent être coordonnées entre diverses sphères de compétences. Ceci est particulièrement vrai en C.-B. et dans le sud-est de l’Alaska, car la plus importante agrégation reproductrice d’otaries de Steller se trouve sur l’île Forrester, à moins de 50 km au nord de la frontière canado-américaine. À la suite d’une recommandation du Steller Recovery Plan (NMFS, 1992) du National Marine Fisheries Service (NMFS), on a consenti des efforts pour normaliser les techniques de recensement (Olesiuk et al.,2008) et pour coordonner les calendriers des évaluations de l’État et de la province (MPO, 2008d), ce qui a donné lieu à la première évaluation exhaustive de la population de l’Est sur l’ensemble de son aire de répartition (Pitcher et al.,2007).
Parmi les principales lacunes dans les connaissances concernant l’otarie de Steller en C.-B., mentionnons la composition du régime alimentaire et les besoins annuels en proies de l’espèce, selon l’âge et la saison. L’abondance et la répartition saisonnières des espèces qui constituent les proies sont également méconnues, tout comme les aires d’alimentation potentiellement importantes. À ce titre, la répartition spatiale et temporelle des pêches peut devenir un facteur de plus en plus important au fur et à mesure que l’on comblera ces lacunes dans les connaissances et que l’on améliorera notre compréhension de l’écologie alimentaire de l’otarie de Steller.
Plusieurs facteurs limitatifs naturels sont actuellement méconnus. Dans certains cas, des programmes de recherche portent sur ces incertitudes, mais aucun résultat n’est actuellement disponible ou n’apporte de réponses concrètes. On a besoin d’information sur les indices vitaux clés de l’otarie de Steller pour déterminer les taux de survie et la fécondité selon l’âge et le sexe, l’âge au sevrage et l’âge à la première reproduction, facteurs qui régissent la productivité de la population. La collecte d’autres échantillons génétiques de la population de la C.-B. pourra améliorer notre connaissance de la répartition entre les roqueries et de la cartographie génétique des individus qui recolonisent la roquerie du groupe Sea Otter.
On ne connaît pas précisément les changements saisonniers du régime alimentaire ou des besoins en matière de proies de la population d’otaries de Steller de la C.-B. L’incertitude est particulièrement importante quant à son régime alimentaire en dehors de la saison de reproduction. Plusieurs études sur le régime alimentaire d’été des otaries de Steller indiquent que les poissons fourrages, comme le hareng, le lançon et la sardine, ainsi que d’autres poissons de taille moyenne vivant en bancs tels que le saumon, la merluche et le sébaste (voir la section « Besoins en matière de proies ») peuvent constituer des composants importants du régime alimentaire (Pike, 1958; Spalding, 1964; Olesiuk et Bigg, 1988, Trites et Olesiuk, données non publiées). D’autres études sur la répartition de ces proies nous aideront à cibler les zones d’alimentation géographiques importantes (en particulier à proximité des roqueries) ainsi que les zones géographiques et temporelles dans lesquelles des interactions avec les pêches sont possibles.
L’importance de la prédation exercée par les épaulards en tant que facteur limitatif demeure quelque peu incertaine. Même si la prédation par les épaulards migrateurs est importante, l’importance spécifique du sexe et de la classe d’âge des otaries qui figurent dans le régime alimentaire de ces épaulards demeure inconnue. L’accroissement de nos connaissances sur la répartition saisonnière des épaulards migrateurs et de leur régime alimentaire nous aidera à déterminer la mesure dans laquelle la prédation gouverne la croissance de la population d’otaries de Steller en C.-B.
Les menaces anthropiques ayant une incidence sur les otaries de Steller en Amérique du Nord et pour lesquelles il faut obtenir davantage d’éclaircissements sont les suivantes :
- Les effets que les activités de recherche ont sur la survie des petits, comme la perturbation aux échoueries et, en particulier, aux roqueries ainsi que le marquage au fer rouge des petits.
- Les concentrations de référence des polluants chimiques et biologiques dans la population de la C.-B.
- L’importance de certains engins de pêche et installations aquicoles (p. ex. engins de traîne vs sennes) en ce qui concerne le risque d’emmêlements.
- La répartition spatiale et temporelle des pêches par rapport aux importantes échoueries et roqueries d’otaries ainsi que la fréquence des interactions avec les pêches, de l’abattage illégal ou des emmêlements.
Le présent plan de gestion a deux buts.
- Faire en sorte que les menaces anthropiques d’origine canadienne ne provoquent pas un déclin insoutenable de la population ou une contraction de l’aire de répartition ou du nombre de sites de reproduction que l’on observe actuellement au Canada.
- Assurer un soutien et une contribution à un environnement dans lequel les activités de recherche et de surveillance concernant les otaries de Steller en C.-B. contribuent à l’amélioration de notre compréhension d’ensemble de la population de l’est du Pacifique.
La population d’otaries de Steller de l’est du Pacifique continue de croître, et aucun signe n’indique qu’elle a atteint la capacité biotique. On peut donc s’attendre que la croissance de la population de cette espèce dépendante des proies se stabilise à un moment donné et fluctue en fonction de changements naturels survenant dans l’abondance des proies. Le rôle des gestionnaires canadiens chargés de cette espèce est de protéger la population vivant dans les eaux canadiennes des sources anthropiques de mortalité qui mettraient en péril la durabilité de cette ressource.
Comme il existe des lacunes importantes dans notre compréhension de l’écologie de cette espèce, de la dynamique de ses populations et des impacts des menaces connues, il est essentiel, pour atteindre cet objectif de gestion, de combler les lacunes dans les connaissances. L’espèce affiche une répartition transfrontalière, et des travaux de recherche ainsi que des initiatives de gestion sont actuellement menés par les gouvernements canadien et américain ainsi que par des organismes privés et des chercheurs indépendants. Le présent plan de gestion reconnaît que les initiatives de gestion et les travaux de recherche sont menés d’une manière plus efficace lorsque toutes les parties coordonnent leurs efforts et collaborent, lorsque c’est possible.
Objectifs relatifs à la population
P1 Maintenir une population viable et éviter que celle-ci ne décline pour atteindre des effectifs auxquels elle pourrait être considérée comme étant à risque d’extinction.
Objectifs relatifs à la répartition
D1 Maintenir l’utilisation annuelle des trois principales roqueries et soutenir la conversion du quatrième site de reproduction en une roquerie permanente.
D2 Maintenir le nombre d’échoueries en Colombie-Britannique ainsi que leur utilisation afin d’assurer une répartition étendue de l’espèce sur toute la côte de la C.-B.
Le maintien de l’abondance et de la répartition des otaries de Steller en C.-B. au cours des trois prochaines générations nous permettra de protéger cette population au Canada. Les échoueries et les roqueries connues, si elles sont protégées, fourniront l’habitat terrestre requis sur l’ensemble de la côte. Comme la population affiche une tendance soutenue à la hausse depuis plus de quatre décennies et que ses effectifs sont supérieurs aux pics historiques connus, les données dérivées des activités de surveillance nous aideront à détecter toute tendance à la baisse au sein de la population ou toute contraction de son aire de répartition.
Objectifs en matière de recherche et de surveillance
Les objectifs en matière de recherche et de surveillance exposés dans le présent plan de gestion sont axés sur les besoins prioritaires suivants.
R1 Mener des évaluations de la population dans l’ensemble de son aire de répartition par l’entremise de relevés coordonnés entre le Canada et les États-Unis, lorsque c’est possible.
R2 Contribuer à accroître et à renforcer notre compréhension de la biologie et des besoins en matière d’habitat de l’otarie de Steller en C.-B.
R3 Soutenir et favoriser la recherche visant à combler les lacunes dans les connaissances relatives aux menaces relevées (tableau 1) et à d’autres menaces éventuelles pesant sur cette population, et contribuer à cette recherche.
R4 Établir les taux cumulatifs totaux de mortalité annuelle d’origine anthropique qui peuvent être maintenus chez les otaries de Steller de la C.-B.
La recherche doit être axée sur les principales lacunes dans les connaissances relatives à cette espèce qui sont précisées dans la section 1.7, « Lacunes dans les connaissances », ainsi que sur les lacunes concernant les effets des menaces énumérées à la section 1.5 du présent plan de gestion.
À l’heure actuelle, les relevés de la population présente dans les eaux de la C.-B. sont d’ordinaire menés tous les quatre ans. Comme les otaries de Steller occupent une aire de répartition qui franchit les limites nord et sud de la frontière canadienne, il faut mettre l’accent sur la coordination de ces relevés si l’on veut obtenir une estimation globale de la population d’otaries de Steller de l’est du Pacifique. Cela nous permettra, en bout de ligne, d’accroître notre compréhension de l’importance relative du composant canadien de cette population. Une surveillance plus fréquente, en particulier aux sites de reproduction locaux, améliorera notre capacité à détecter les déclins démographiques et à intervenir.
On a établi que la recherche sur l’écologie de l’alimentation, y compris l’évaluation des interactions possibles avec les pêches, constituait une priorité au Canada. En outre, des programmes pour préciser les indices vitaux, l’utilisation de l’habitat et la cartographie génétique à échelle réduite nous aideront à orienter la gestion et la protection de cette population dans les eaux de la C.-B.
L’otarie de Steller est vulnérable à un certain nombre de sources de mortalité anthropique. Nous devons mieux comprendre les taux actuels de mortalité anthropique et leurs tendances au fil du temps. L’établissement de seuils durables pour la mortalité d’origine anthropique (objectif R4) permettra la surveillance des effets au niveau de la population. Les populations d’otaries de Steller se sont accrues au cours des dernières années, ce qui indique que les taux actuels de mortalité anthropique ne mettent pas en péril la durabilité de l’espèce. Cependant, la situation pourrait changer si les conditions devenaient moins favorables pour les otaries, auquel cas la mortalité pourrait réduire les taux de croissance de la population ou accroître l’ampleur des fluctuations démographiques.
Objectifs de gestion
Le tableau 1 (section 1.5.1) résume l’évaluation des menaces pesant sur les otaries de Steller de l’est du Pacifique et présente le potentiel d’atténuation perçu pour chaque menace. L’analyse de la gravité, de l’occurrence et de la certitude causale a permis d’établir l’importance globale de chaque menace relativement aux impacts qu’elle peut avoir sur la viabilité de cette population. Au cours des dix prochaines années, les objectifs de gestion suivants nous aideront à faire en sorte que les menaces les plus importantes n’aient pas d’incidence sur la population d’otaries de Steller ou sur sa répartition en C.-B. et à contribuer à la conservation de cette population dans l’ensemble de son aire de répartition.
M1 Promouvoir la collaboration internationale, la recherche indépendante, la formation et la vulgarisation concernant les initiatives de gestion et de conservation.
M2 Limiter l’exposition des otaries de Steller aux polluants.
M3 Limiter la perturbation des otaries de Steller aux roqueries pendant la saison de reproduction.
M4 Réduire le risque de déversements catastrophiques ayant une incidence sur les otaries de Steller ou leur habitat au Canada.
M5 Limiter l’exposition des otaries de Steller à des bruits aigus qui provoquent des perturbations comportementales ou physiques chez les pinnipèdes.
M6 Limiter la probabilité que les proies soient limitées par des facteurs anthropiques.
La population affiche une tendance soutenue à la hausse. Le but des objectifs de gestion portant sur les menaces les plus importantes est d’éviter toute diminution non souhaitée de la population et de prévenir la survenue de changements importants dans l’aire de répartition de l’espèce dans les eaux de la C.-B. Les déversements catastrophiques, les perturbations (telles que les perturbations acoustiques, celles découlant de l’exploitation des ressources énergétiques et minérales ainsi que les perturbations physiques aux roqueries) et les contaminants sont considérés comme étant des menaces anthropiques qui doivent faire l’objet de mesures d’atténuation ou de gestion afin que l’on puisse atteindre le but visé dans le présent plan de gestion. La limitation de l’abondance des espèces-proies par des moyens anthropiques (obj. M5) est d’une importance particulière; cependant, on manque d’information sur les besoins en matière de proies. Ainsi, la surveillance de la disponibilité des proies et l’exécution de travaux de recherche nous aideront à prévenir les impacts sur la population.
Les menaces qui ont été évaluées comme étant d’importance faible ou inconnue ne font pas l’objet d’objectifs particuliers ni de nouvelles recommandations en matière d’atténuation; les lacunes dans les connaissances se rapportant à ces menaces seront plutôt comblées par des moyens opportunistes ou peu coûteux lorsque cela sera possible. Certaines menaces ont une incidence sur les individus, mais n’affectent pas l’ensemble de la population. Lorsque la possibilité de mettre en œuvre des mesures d’atténuation efficaces est élevée (tableau 1) et que des ressources sont disponibles, il est prudent de gérer ces menaces et d’atténuer leurs effets.
Les mesures suivantes (non classées par ordre de priorité) soutiennent les objectifs exposés à la section 2.2. Certaines des mesures énumérées ci-après sont en cours (voir section 1.6, « Mesures déjà prises ou en cours ») et ont été décrites dans d’autres documents de planification du rétablissement jusqu’à maintenant (voir section 4.0, « Plans connexes »). La synchronisation des activités énumérées pour la protection, la gestion et la recherche facilitera l’application d’une approche plurispécifique à la conservation des mammifères marins en C.-B. et permettra une utilisation efficace des ressources disponibles. Les mesures sont recommandées lorsque leur mise en œuvre est considérée comme étant réalisable et que celle-ci aura lieu là où elles seront vraisemblablement les plus efficaces pour assurer la protection de la population d’otaries de Steller en C.-B.
Lorsque les mesures à prendre relèvent de la compétence de Pêches et Océans Canada ou de l’Agence Parcs Canada, celles-ci seront mises en œuvre directement sous réserve de la disponibilité des fonds et des autres ressources requises. La collaboration d’autres organismes et agences responsables sera cependant nécessaire pour la mise en œuvre de certaines mesures. Lorsque la responsabilité de la mise en œuvre des mesures ne relève pas du mandat ou de la compétence de Pêches et Océans Canada ou de l’Agence Parcs Canada, il sera prioritaire de soutenir la mise en œuvre de la ou des mesures et la contribution aux efforts consentis, lorsque cela est possible. Les agences participantes, les organismes et les échéanciers de mise en œuvre pour chacune des mesures énumérées sont présentés à la section 3 (tableau 3). Les organismes qui participent actuellement à la collecte de données sur la population d’otaries de Steller de l’est du Pacifique sont énumérés à l’annexe II.
1. Protéger les otaries de Steller contre les perturbations (physiques et acoustiques) aux quatre sites de reproduction ou à proximité de ceux-ci, aux 23 échoueries permanentes et à de nombreuses échoueries d’hiver en C.-B.
- Poursuivre la mise en application du Règlement sur les mammifères marins (RMM) et la promotion des lignes directrices régionales sur l’observation des mammifères marins ainsi que des règlements pertinents s’appliquant au développement industriel marin.
2. Poursuivre la mise en œuvre de la réglementation concernant les rejets de débris5 et élaborer de nouvelles normes sur les engins de pêche (engins de pêche à la traîne, sennes et palangres) afin de réduire le risque d’emmêlement dans des débris marins ou d’emmêlement direct dans des engins de pêche.
3. Poursuivre la gestion des ressources halieutiques et des pêches lorsqu’un lien existe avec le régime alimentaire des otaries de Steller. Examiner les besoins alimentaires des otaries de Steller lorsque des changements surviennent dans les régimes de gestion des pêches en vigueur pour les espèces que l’on sait importantes pour l’otarie de Steller en C.-B.
4. Poursuivre l’examen de propositions de projets qui comportent des activités ayant la possibilité de perturber les otaries aux échoueries ou aux roqueries et formuler des avis propres aux projets concernant des mesures d’atténuation ou d’évitement qui tiennent compte des besoins en matière d’habitat des otaries de Steller.
5. Même si les contaminants sont considérés comme une menace pour cette population, la gestion de la contamination biologique et chimique relève de la compétence d’Environnement Canada. Les mesures suivantes sont recommandées pour réduire la charge de substances toxiques chez les mammifères marins et dans leur habitat.
- Élaborer un plan d’intervention d’urgence pour préciser les champs d’expertise en mammifères marins nécessaires pour mener à bien des initiatives d’intervention en cas de déversement, au besoin.
- Élaborer un manuel opérationnel propre aux mammifères marins qui sera inclus dans les plans d’intervention en cas de déversement catastrophique3 afin de préciser les protocoles d’intervention à suivre et les données à recueillir pour assurer l’atténuation et le suivi des effets à court et à long termes sur les mammifères marins et l’habitat important.
- Passer en revue la gestion des sources ponctuelles de polluants toxiques dans l’habitat connu de l’otarie de Steller afin d’évaluer le respect des lignes directrices fédérales, provinciales et régionales concernant les seuils de contamination environnementale établis pour certaines toxines figurant à l’annexe I.
6. Soutenir les activités de recherche à faible impact par la délivrance de permis et la coordination, pour toute l’aire de répartition de l’espèce, de recherches transfrontalières ainsi que les activités de surveillance et d’évaluation (sections 2.3.3. et 2.3.4.) afin de combler les lacunes importantes dans les connaissances et de clarifier les menaces relevées, tout en limitant la perturbation des animaux. On pourra ainsi effectuer une collecte de données uniforme et normalisée pour les besoins des évaluations et des comparaisons des populations et sur l’ensemble de l’aire de répartition.
7. Renforcer les mesures mises en œuvre pour réduire le risque d’emmêlement des otaries de Steller dans des débris marins, des installations aquicoles et des engins de pêche.
- Soutenir le Marine Mammal Response Network (MMRN) dans la collecte de données à l’appui d’une étude approfondie des menaces suivantes : prises accessoires, emmêlement dans des débris marins, des engins de pêche et des installations aquicoles.
- Améliorer, soutenir et favoriser, lorsque c’est possible, l’élaboration continue de normes sur les déclarations par les observateurs des pêches et de lignes directrices pour l’identification des espèces de mammifères marins et la collecte de données afin de préciser l’étendue des interactions avec les pêches pour ce qui est des emmêlements, des prises accessoires ou de la déprédation et de recueillir des exemples, lorsque c’est possible et nécessaire.
- Poursuivre l’élaboration de solutions au problème des interactions avec les prédateurs et pour la déclaration trimestrielle des abattages de mammifères marins aux sites aquicoles, et poursuivre l’élaboration d’exigences en matière de déclaration des interactions avec des mammifères marins aux sites aquicoles. Il est particulièrement important que l’on puisse identifier les espèces avec précision, recueillir des données et établir des exigences en matière de délais pour la déclaration des incidents.
- Poursuivre les travaux engagés avec l’industrie aquicole afin d’évaluer et d’élaborer des pratiques de gestion optimales pour atténuer la déprédation par les otaries de Steller et éviter les emmêlements subséquents (p. ex. infrastructure de filets anti-prédation plus efficace).
- Proposer d’autres types d’engins afin de limiter de façon préventive la probabilité d’emmêlement dans des débris marins, des engins de pêche et des installations aquicoles. Il est particulièrement important que l’on examine le risque d’emmêlement dans les filets anti-prédation et les bandes d’emballage ainsi que l’ingestion d’hameçons de lignes à la traîne et de palangres. Cette mesure contribuera à l’évolution continue des lignes directrices, des pratiques de gestion optimales, des règlements et des normes.
8. Évaluer le risque que font peser sur les otaries de Steller de la C.-B. les déversements catastrophiques et les perturbations attribuables aux bruits et aux navires qui pourraient découler de la levée du moratoire sur la prospection et l’extraction de combustibles fossiles au large de la C.-B. Les impacts sur le succès de l’alimentation en mer près des échoueries permanentes et des sites de reproduction de la C.-B. sont particulièrement préoccupants.
Les domaines suivants ont été identifiés comme étant prioritaires sur le plan des recherches à effectuer pour combler les lacunes entourant la biologie de l’espèce. D’autres domaines de recherche potentiels ont été énumérés dans les sections précédentes du présent plan de gestion et doivent également être pris en considération du fait qu’ils peuvent être utiles pour la mise en œuvre des mesures ci-après.
9. Pêches et Océans Canada soutiendra, facilitera et favorisera la recherche sur les otaries de Steller en assurant la gestion des échantillons et des données recueillies par des organismes non gouvernementaux et le public, notamment :
- Les échantillons de tissus provenant d’individus de la population d’otaries de Steller de la C.-B. pour d’éventuelles recherches sur la génétique, les agents pathogènes et les acides gras. On facilitera ainsi la collecte de données sur la génétique et les agents pathogènes afin que l’on puisse mieux comprendre les exigences et la santé de la population de la C.-B.; on facilitera ainsi également les comparaisons menées sur l’ensemble de l’aire de répartition de l’espèce.
- Les nouvelles données d’observation des marques et d’éventuelles photographies des animaux marqués. On contribuera ainsi à résoudre les incertitudes relatives aux indices vitaux et aux déplacements des individus et à combler d’autres lacunes dans les connaissances.
10. Pêches et Océans Canada et l’Agence Parcs Canada soutiendront, faciliteront et favoriseront, lorsque c’est possible, la mise en œuvre des mesures de recherche dirigées suivantes afin de combler les principales lacunes dans les connaissances relatives aux otaries de Steller.
- Études sur les aires d’alimentation et la répartition saisonnière des otaries ainsi que de leurs proies afin de résoudre les incertitudes concernant la disponibilité des proies près des roqueries.
- Études afin de résoudre les incertitudes concernant le régime alimentaire des otaries de Steller en dehors de la saison de reproduction.
- Tenue d’une base de données sur les nouvelles observations d’individus marqués afin de faciliter l’organisation des données recueillies et le partage des données entre les chercheurs.
Pêches et Océans Canada et l’Agence Parcs Canada soutiendront, faciliteront et favoriseront, lorsque c’est possible, la mise en œuvre des mesures de recherche suivantes afin de combler les lacunes dans les connaissances relatives aux menaces pesant sur la population.
11. Évaluer les taux actuels de mortalité anthropique annuelle qui peuvent être soutenus par les otaries de Steller dans diverses conditions (tout en atteignant l’objectif P1) ainsi que les tendances connexes.
12. Déterminer la composition du régime alimentaire des otaries de Steller (pour toutes les saisons) afin d’évaluer les possibilités de concurrence avec les pêches à l’égard des ressources que représentent les proies.
13. Évaluer la variation saisonnière de l’importance des impacts des perturbations causées par les activités de recherche menées aux échoueries et aux roqueries afin de clarifier l’ampleur de cette menace relevée.
14. Recueillir de l’information sur les taux d’emmêlement observés pendant les recherches sur l’otarie de Steller en C.-B. afin de compiler de l’information sur les taux d’emmêlement, les types d’engins et la gravité des emmêlements ainsi que d’examiner et d’élaborer des méthodes pour libérer les animaux emmêlés.
15. Recueillir des échantillons à des fins d’analyse des charges en toxines et en agents pathogènes chez les otaries de Steller, lorsque cela est possible.
16. Procéder à des évaluations des sources actuelles de polluants biologiques dans l’habitat connu de l’otarie de Steller afin d’évaluer la possibilité d’impacts sur toute la population. Une importance particulière est accordée aux sources agricoles de polluants et aux programmes de réhabilitation.
17. Identifier les sources diffuses importantes de contaminants toxiques dans l’habitat de l’otarie de Steller en C.-B. Une importance particulière est accordée aux contaminants nouveaux et aux sources situées à proximité immédiate des roqueries et des échoueries permanentes.
18. Continuer de soutenir et de faciliter la coordination de relevés aériens et par navire menés tous les quatre ans dans l’ensemble de l’aire de répartition.
19. Envisager l’exécution de relevés supplémentaires aux deux ans aux roqueries afin d’assurer un suivi plus étroit des populations reproductrices.
20. Favoriser l’amélioration des réseaux de communication afin d’accroître la sensibilisation à l’égard des initiatives de conservation de l’otarie de Steller et de la recherche menée à cet égard.
- Constituer, dans le cadre des activités de prévention, des réseaux intra et interagence afin d’assurer une communication efficace pendant les interventions en cas de déversement catastrophique et permettre la prise de mesures rapides, efficaces et coordonnées par les agences et parties responsables.
- Soutenir et faciliter, lorsque c’est possible, la collaboration transfrontalière et intergouvernementale concernant des initiatives de conservation afin d’assurer une réaction coordonnée pour la conservation de cette population.
- Communiquer de façon continue avec les médias et faire la promotion du RMM ainsi que du document « Respectez les baleines! Directives pour l’observation de la faune aquatique à l’intention des plaisanciers et des observateurs » afin de réduire la perturbation physique et acoustique aux échoueries. Cette mesure contribuera à accroître la sensibilisation des gens au Règlement et facilitera la mise en œuvre de mesures d’application de la réglementation.
- Soutenir, dans la mesure du possible, les programmes d’éducation et de sensibilisation indépendants sur les sujets suivants.
- Importance des cas d’emmêlement dans des débris marins observés chez des otaries de Steller.
- Mesures pour éviter ou limiter la déprédation dans les engins de pêches commerciale et sportive.
- Importance de déclarer les incidents mettant en cause des mammifères marins, y compris l’otarie de Steller, au MMRN.
- Mesures d’atténuation disponibles pour réduire la contamination chimique et biologique de l’habitat.
Pêches et Océans Canada et l’Agence Parcs Canada encouragent d’autres agences et organismes à prendre part aux activités de conservation de l’otarie de Steller, population de l’est du Pacifique, par l’entremise du présent plan de gestion. Les agences et organismes mentionnés au tableau 2 sont des partenaires possibles pour la mise en œuvre des mesures recommandées.
Le tableau 3 résume les mesures recommandées pour soutenir les buts et les objectifs du présent plan de gestion. Les activités mises en œuvre par Pêches et Océans Canada sont conditionnelles à la disponibilité des fonds et des autres ressources nécessaires. Au besoin, des partenariats seront conclus avec des organismes et des secteurs particuliers afin que l’on dispose de l’expertise et des ressources requises pour prendre les mesures énumérées. Toutefois, la liste des partenaires n’est présentée qu’à titre indicatif pour les autres organismes, et l’exécution des mesures en question dépendra des priorités et des contraintes budgétaires de chaque agence ou organisme. Les agences ou organismes qui recueillent actuellement des données sur l’otarie de Steller sont énumérés à l’annexe II.
Tableau 2 . Les mesures de gestion indiquées dans le présent plan doivent être mises en œuvre, lorsque cela est approprié, en partenariat avec les organismes suivants.
| Organisation | Acronyme |
|---|---|
| Pêches et Océans Canada | MPO |
| Agence Parcs Canada | APC |
| Marine Mammal Response Network | MMRN |
| Ministère de la Défense nationale | MDN |
| Environnement Canada | EC |
| Transports Canada | TC |
| Ressources naturelles Canada | RNCan |
| Garde côtière canadienne | GCC |
| Office national de l’énergie | ONE |
| Premières nations | PN |
| Gouvernement de la Colombie-Britannique | Gouv. de la C.-B. |
| Ministère de l’Agriculture et des Terres | MAL |
| Vancouver Aquarium Marine Science Centre | VAMSC |
| B.C. Cetacean Sightings Network | B.C. CSN |
| Commission du saumon du Pacifique | CSP |
| Straitwatch | Straitwatch |
| Universités menant des programmes de recherche pertinents | Universités |
| U.S. National Marine Fisheries Service | NMFS |
| National Oceanic & Atmospheric Administration, National Marine Mammal Lab |
NOAA |
| Oregon Department of Fish and Wildlife | ODFW |
| Washington Department of Fish and Wildlife | WDFW |
| Alaska Department of Fish and Game | ADFG |
| Whale Watch Operators Association Northwest | WWOANW |
| Organisations non gouvernementales de l’environnement | ONGE |
| North Pacific Marine Mammal University Consortium | NPMMUC |
| À déterminer | – |
Tableau 3. Calendrier de mise en œuvre proposé
Mesure |
Obj. | Priorité | Menaces ou préoccupations étudiées | Agences/organismes participants4 | Échéancier | ||||
| Protection | |||||||||
| 1. Protéger l’otarie de Steller contre les perturbations aux sites de reproduction et aux échoueries. | |||||||||
| a. Poursuivre la mise en application du RMM et promouvoir les lignes directrices régionales | D1, D2, M3 |
F | Protection continue contre les perturbations; dégradation de l’habitat | GCC, MPO, APC | En cours | ||||
| 2. Mettre en œuvre la réglementation concernant les rejets de débris; élaborer de nouvelles normes sur la pêche | M2 | F | Réduction du risque d’emmêlement | MPO, TC, industrie de la pêche | En cours | ||||
| Gestion | |||||||||
| 3. Examiner les besoins alimentaires lorsque des changements surviennent dans les régimes de gestion des pêches en vigueur pour les espèces que l’on sait importantes pour l’otarie de Steller en C.-B. ou qui pourraient le devenir | P1, M6 | É | Limitation des proies disponibles | MPO, industrie de la pêche | Lorsque des changements sont apportés à la gestion des pêches | ||||
| 4. Examiner les propositions pouvant entraîner une perturbation aux échoueries et aux roqueries et formuler des avis | D1, D2, M3, M5 |
F | Perturbation par les bruits sismiques; utilisation de sonars tactiques | MPO, MDN, industrie, RNCan, ONE | En cours, accroître la participation au besoin | ||||
| 5. Gérer et réduire les apports de toxines chimiques, réduire la charge en substances toxiques | |||||||||
| a. Élaborer un plan d’intervention d’urgence pour préciser les champs d’expertise en mammifères marins nécessaires pour mener à bien des initiatives d’intervention en cas de déversement de substances toxiques | P1, M2, M4 |
FM | Intervention efficace et coordonnée en cas de déversement de substances toxiques qui peuvent affecter les proies ou l’habitat de l’otarie de Steller | MPO, EC, GCC, APC, gouv. de la C.-B., TC | 4 ans | ||||
| b. Élaborer un manuel opérationnel propre aux mammifères marins portant sur l’intervention en cas de déversement de substances toxiques | M2, M4 | FM | Intervention progressive efficace et coordonnée en cas de déversement de substances toxiques; collecte de données normalisée; surveillance | MPO, EC, GCC, APC, gouv. de la C.-B., TC | 4 ans | ||||
| c. Passer en revue la gestion des sources ponctuelles de toxines figurant à l’annexe I | R3, M2 | FM | Utilité des lignes directrices et des seuils à l’égard des effets physiologiques sur les mammifères marins occasionnés par la charge en contaminants dans l’habitat et chez les proies | EC, Gouv. de la C.-B., municipalités, industrie | En cours | ||||
| 6. Soutenir la recherche à faible impact par la délivrance de permis et la coordination de programmes de recherche | R1 à R4, M1, M3 | É | Promotion de la recherche à faible impact indépendante ou menée par des entités autres que le MPO, de la collaboration transfrontalière et du partage des données | MPO, ONGE, NPMMUC, NMFS, ADFG, WDFW, ODFW, à déterminer |
En cours | ||||
| 7. Renforcer les mesures mises en œuvre pour réduire le risque d’emmêlement | |||||||||
| a. Soutenir les initiatives du MMRN | R3, R4 | F | Collecte de données sur les prises accessoires, les emmêlements (de tous types) et les prises accidentelles | MPO, VAMSC, ONGE, MAL, NPMMUC, APC, GCC, à déterminer | En cours | ||||
| b. Poursuivre l’élaboration de normes et de lignes directrices sur les déclarations par les observateurs des pêches | R3, R4 | M | Clarification de l’ampleur des interactions avec les pêches | MPO, industrie de la pêche | 3 ans | ||||
| c. Poursuivre l’élaboration de solutions au problème des interactions avec les prédateurs et la déclaration trimestrielle sur les abattages de mammifères marins aux sites aquicoles | P1, R3, R4 | F | Clarification de l’ampleur des interactions aux sites aquicoles; emmêlement, lutte contre les prédateurs | MPO, industrie de l’aquaculture | En cours | ||||
| d. Poursuivre les travaux engagés avec l’industrie aquicole afin d’évaluer et d’élaborer des pratiques de gestion optimales | P1, M2 | F | Atténuation de la déprédation et des emmêlements | MPO, industrie de l’aquaculture | En cours | ||||
| e. Proposer d’autres types d’engins ou d’installations (pêche, aquaculture) | P1, M2 | M | Réduction du risque d’emmêlement; prises accidentelles | MPO, industries de la pêche et de l’aqua-culture, VAMSC | En cours; lorsque de nouvelles informations sur les engins ou les installations seront disponibles | ||||
| 8. Évaluer le risque accru qui pourrait découler de la levée du moratoire sur l’extraction de combustibles fossiles au large | P1, D1, D2, R3, M2 à M5 |
FM | Détermination du risque associé aux déversements de substances toxiques; la perturbation; les impacts potentiels sur l’alimentation | MPO, EC, ONE, RNCan, gouv. de la C.-B. | En cours | ||||
| Recherche sur la biologie de l’otarie de Steller | |||||||||
| 9. Gérer les échantillons et les données recueillies sur les nouvelles observations d’individus marqués. | |||||||||
| a. Prélever des échantillons de tissus | R2, R3, M1 | F | Analyses génétiques, analyses des agents pathogènes et des acides gras | MPO, NMFS, ODFW, ADFG, WDFW, à déterminer | Sur une base opportuniste | ||||
| b. Observer de nouveau et photographier des animaux marqués | R1, R2, R3, M1 | M | Contribution aux efforts de recherche sur l’ensemble de l’aire de répartition afin d’évaluer les indices vitaux et les déplacements | MPO, NMFS, ODFW, ADFG, WDFW, à déterminer | En cours | ||||
| 10. Soutenir, faciliter et favoriser la recherche dans les domaines suivants. | |||||||||
| a. Études sur les aires d’alimentation importantes et la répartition saisonnière | P1, R2, R3, M1, M6 |
É | Disponibilité des proies près des roqueries pendant la période de reproduction | MPO, CSP, WDFW, ADFGW, NMFS | En cours | ||||
| b. Études sur le régime alimentaire en dehors de la saison de reproduction | P1, R2, R3, M1, M6 | É | Lacunes dans les connaissances : espèces-proies importantes sur une base saisonnière; besoins nutritionnels | MPO, CSP, WDFW, NPMMUC, NMFS, ODFW, à déterminer | En cours | ||||
| c. Tenue d’une base de données sur les nouvelles observations d’individus marqués | R2, M1 | M | Partage des données; organisation des données | MPO, ADFGW, ODFW, NMFS, NPMMUC | En cours | ||||
| Recherche pour clarifier les menaces relevées | |||||||||
| 11. Évaluer la durabilité de la mortalité anthropique totale | P1, R4 | M | Taux durables de mortalité anthropique (permettant l’atteinte de l’objectif P1) | MPO, à déterminer | En cours | ||||
| 12. Évaluer les possibilités de concurrence avec les pêches à l’égard des ressources que représentent les proies (pour toutes les saisons) | P1, R3, M6 |
É | Limitation des proies disponibles; impacts des pêches | MPO, CSP, WDFW, NPMMUC | En cours | ||||
| 13. Déterminer la variation saisonnière de l’importance de la perturbation causée par les activités de recherche menées aux échoueries et aux roqueries | D1, D2, R3, M3 |
FM | Importance des perturbations saisonnières | NPMMUC, à déterminer | En cours | ||||
| 14. Recueillir de l’information sur les taux d’emmêlement observés pendant les recherches | P1, R3, M1, M2 |
F | Taux d’emmêlement en C.-B.; types d’engins, gravité des incidents | MPO, NMFS, ODFW, ADFG, WDFW, VAMSC, NPMMUC, à déterminer | En cours | ||||
| 15. Analyser les charges en toxines et en agents pathogènes | P1, R3, M2 |
FM | Détermination du risque d’impacts associés aux toxines et aux agents pathogènes | MPO, EC, gouv. de la C.-B., MAL, à déterminer | En cours | ||||
| 16. Évaluer les sources de polluants biologiques | R3, M2 | F | Détermination du potentiel associé à la charge de polluants biologiques | Municipalités, industrie, à déterminer | En cours | ||||
| 17. Identifier les sources diffuses importantes de contaminants toxiques | R3, M2 | FM | Nouveaux contaminants; charges en substances toxiques à proximité des roqueries et des échoueries permanentes | À déterminer | En cours | ||||
| Suivi de l’état de la population | |||||||||
| 18. Soutenir et faciliter la coordination de relevés menés tous les quatre ans dans l’ensemble de l’aire de répartition | P1, D1, D2, R1, M1 |
É | Suivi des tendances dans l’ensemble de l’aire de répartition; collecte de données efficace; promotion du partage des données | MPO, APC, gouv. de la C.-B., NMFS, ODFW, ADFG, WDFW, à déterminer | En cours (prochain relevé en 2010) |
||||
| 19. Envisager l’exécution de relevés aux deux ans aux roqueries | P1, D1, D2, R1, R2, M1 |
M | Suivi des populations reproductrices | MPO, APC, gouv. de la C.-B., NMFS, ODFW, ADFG, WDFW, à déterminer | Effectué en 2008; le prochain relevé sera fonction du financement disponible | ||||
| Vulgarisation et communication | |||||||||
| 20. Favoriser l’amélioration des réseaux de communication | |||||||||
| a. Constituer, dans le cadre des activités de prévention, des réseaux de communication | M1,M2, | M | Communication intra et interagence efficace en cas de déversement catastrophique | MPO, EC, GCC, gouv. de la C.-B., VAMSC, municipalités, ONGE | 3 ans | ||||
| b. Soutenir et faciliter, dans la mesure du possible, les initiatives de collaboration transfrontalière et intergouvernementale | Tous | M | Gestion et efforts de recherche coordonnés | MPO, APC, NOAA, NMFS, NPMMUC, ONGE | 3 ans | ||||
| c. Communiquer de façon continue avec les médias et faire la promotion du RMM ainsi que du document « Respectez les baleines! » | M1 | F | Sensibilisation aux lignes directrices, au Règlement; mesures d’application de la réglementation | MPO, B.C. CSN, VAMSC, Straitwatch, WWOANW, à déterminer | En cours | ||||
| d. Soutenir, dans la mesure du possible, des programmes d’éducation et de sensibilisation indépendants | M1, M2 | F | Emmêlement; débris marins; déprédation; déclaration des incidents; contamination de l’habitat | MPO, B.C. CSN, VAMSC, Straitwatch, WWOANW, à déterminer | En cours | ||||
Les documents dont il est question ci-après sont des plans ou des programmes de rétablissement qui relèvent des menaces pesant sur des espèces de mammifères marins en péril et qui contiennent des recommandations à l’égard des mesures d’atténuation de ces menaces, lesquelles sont semblables à celles dont il est question dans le présent plan de gestion de l’otarie de Steller au Canada. La mise en œuvre des mesures mentionnées dans le présent document et dans les plans et programmes de rétablissement énumérés ci-après suivra une approche plurispécifique et plurigouvernementale axée sur la conservation des mammifères marins sur la côte ouest de l’Amérique du Nord.
- U.S. National Marine Fisheries Service. Draft revised Recovery Plan for the Steller sea lion (Eumetopias jubatus) (NMFS, 2007).
- Plan de gestion de l’épaulard du large (Orcinus orca) au Canada [Proposition] (MPO, 2008a).
- Plan de gestion (version provisoire) de la baleine grise (Eschrichtius robustus) dans les eaux canadiennes du Pacifique (MPO, 2008b).
- Plan de gestion du marsouin commun (Phocoena phocoena) du Pacifique au Canada [Proposition] (MPO, 2008c).
- Programme de rétablissement de l’épaulard migrateur (Orcinus orca) au Canada [Version finale] (MPO, 2007c).
- Programme de rétablissement de l’épaulard résident du Nord et du Sud (Orcinus orca) au Canada [Version finale] (MPO, 2008).
- Programme de rétablissement de la loutre de mer (Enhydra lutris) au Canada [Version finale] (MPO, 2007b).
Addison, R.F. 1989. Organochlorines and marine mammal reproduction. Journal canadien des sciences halieutiques et aquatiques 46: 360-368.
Ainley, D.G., C.S. Strong, H.R. Huber, T.J. Lewis et S.H. Morrell. 1981. Predation by sharks on pinnipeds at the Farallon Islands. Fishery Bulletin 78: 941-945.
Alaska Sea Grant. 1993. Is it food? Addressing marine mammal and sea bird declines. Alaska Sea Grant AK-SC-93-01. 65 p.
Alverson, D. 1992. A review of commercial fisheries and the Steller sea lion (Eumetopias jubatus): the conflict arena. Reviews in Aquatic Sciences 6: 203-256.
Anderson, P.J., J.E. Blackburn et B.A. Johnson. 1997. Declines in forage species in the Gulf of Alaska, 1972-95, as indicator of regime shift. p. 531-543 Dans : Forage Fishes in Marine Ecosystems. Alaska Sea Grant AK-SG-97-01. 816 p.
Anderson, P.J. et J.F. Piatt. 1999. Community reorganization in the Gulf of Alaska following ocean climate regime shift. Marine Ecology Progress Series 189: 117-123.
Andrews, R.D. 1999. Preliminary progress report of Steller sea lion (SSL) foraging ecology studies. Présentation au Steller Sea Lion Research Peer Review Feeding Ecology Workshop, Seattle, WA, 11-12 février 1999. 40 p.
Angliss, R.P., D.P. DeMaster et A.L. Lopez. 2001. Alaska Marine Mammal Stock Assessments, 2001. Mémoire technique de la NOAA. NMFS-AFSC-124. 206 p.
Angliss, R.P. et R.B. Outlaw. 2007. Alaska Marine Mammal Stock Assessments, 2006. Mémoire technique de la NOAA, NMFS-AFSC–168. 244 p.
Arndt, D.P. 1973. DDT and PCB levels in three Washington State harbor seal (Phoca vitulina richardsi) populations. Mémoire de maîtrise ès sciences, Université de Washington, Seattle, WA. 65 p.
Azana, C.D.P. 2002. Seasonal variation in the nutrient composition of Alaskan walleye pollock (Theragra chalcogramma) and its effect on the nutritional status of Steller sea lions (Eumetopias jubatus). Mémoire de maîtrise ès sciences, Université de la Colombie-Britannique, Vancouver, C.-B. 104 p.
Baird, R.W. et L.M. Dill. 1995. Occurrence and behaviour of transient killer whales: seasonal and pod-specific variability, foraging behaviour, and prey handling. Revue canadienne de zoologie 73: 1300-1374.
Baker, C.S., T.R. Loughlin, V. Burkanov, C.W. Matson, R.G. Trujillo, D.G. Calkins, J.K. Wickliffe et J.W. Bickham. 2005. Variation of mitochondrial control region sequences of Steller sea lions: the three-stock hypothesis. Journal of Mammalogy 86(6): 1075-1084.
Barrett-Lennard, L.G., K. Heise, E. Saulitis, G. Ellis et C. Matkin. 1995. The impact of killer whale predation on Steller sea lion populations in British Columbia and Alaska. Rapport à l’intention du North Pacific Universities Marine Mammal Research Consortium. Université de la Colombie-Britannique, Vancouver, C.-B. 77 p.
Barron, M.G., R. Heintz et M.M. Krahn. 2003. Contaminant exposure and effects in pinnipeds: implications for Steller sea lion declines in Alaska. Science of the Total Environment 311: 111-133.
Beach, R.J., A.C. Geiger, S.J. Jeffries, S.D. Treacy et B.L. Routman. 1985. Marine mammals and their interactions with fisheries of the Columbia River and adjacent waters, 1980-82. NMFS, NWAFC Processed Report. 85-04. 316 p.
Beamish, R.J. et D.R. Bouillon. 1993. Pacific salmon production trends in relation to climate. Journal canadien des sciences halieutiques et aquatiques 50: 1002-1016.
Benson, A.J. et A.W. Trites. 2002. Ecological effects of regime shifts in the Bering Sea and eastern North Pacific Ocean. Fish and Fisheries 3: 95-113.
Bergman, A., M. Olsson et S.Reiland. 1992. Skull-bone lesions in the Baltic grey seal (Halichoerus grypus). Ambio 21: 517-519.
Beverton, R.J.H. 1985. Analysis of marine mammal-fisheries interaction. p. 3-33. Dans : J.R. Beddington, R.J.H. Beverton et D.M. Lavigne (éd.). Marine Mammals and Fisheries. George Allen and Unwin, Londres, R.-U.
Bickham, J.W. 2000. Variation in nuclear microsatellites and mtDNA of Steller sea lions taken from rookeries and native harvests. Rapport non publié du National Marine Mammal Laboratory. Seattle, WA. 21 p.
Bickham, J.W., J.C. Patton et T.R. Loughlin. 1996. High variability for control-region sequences in a marine mammal: implications for conservation and biogeography of Steller sea lions (Eumetopias jubatus). Journal of Mammalogy 77: 95-108.
Bigg, M.A. 1984. Sighting and kill data for the Steller sea lion (Eumetopias jubatus) and California sea lion (Zalophus californianus) in British Columbia, 1892-1982, with some records from Washington and southeastern Alaska. Rapport canadien des sciences halieutiques et aquatiques 460: 1-191.
Bigg, M.A. 1985. Status of Steller sea lion (Eumetopias jubatus) and California sea lion (Zalophus californianus) in British Columbia. Publication spéciale canadienne des sciences halieutiques et aquatiques 77: 1-20.
Bigg, M.A. 1988. Status of the Steller sea lion Eumetopias jubatus in Canada. Canadian Field-Naturalist 102: 315-336.
Bigg, M.A., G.M. Ellis, P. Cottrell et L. Milette. 1990. Predation by harbour seals and sea lions on adult salmon in Comox Harbour and Cowichan Bay, British Columbia. Rapport technique canadien des sciences halieutiques et aquatiques 1769: 1-31.
Bigg, M.A. et I. Fawcett. 1985. Two biases in diet determination of northern fur seals (Callorhinus ursinus). p. 284-291. Dans : J.R. Beddington, R.J.H. Beverton et D.M. Lavigne (éd.). Marine Mammals and Fisheries. George Allen and Unwin, Londres, R.-U.
Bonnell, M.L., M.O. Pierson et G.D. Farrens. 1983. Pinnipeds and sea otters of central and northern California, 1980-1983: status, abundance and distribution. Final Report, Marine Mammal and Seabird Study, Contract No. 14-12-0001-29090. Prepared by Center for Marine Sciences, University of California, Santa Cruz, for the Pacific OCS Region, Minerals Management Service, OCS Study MMS 84-0044. 220 p.
Bowen, W.D. 1997. Role of marine mammals in aquatic ecosystems. Marine Ecology Progress Series 158: 267-274.
Bowen, W.D., J.I. McMillan et R. Mohn. 2003. Sustained exponential population growth of the grey seal on Sable Island. ICES. Journal of Marine Sciences 60: 1265–1374.
Brenton, C.M. 1977. Inter and intraspecific behaviour of Eumetopias jubatus and Zalophus californianus on a winter haulout area. Mémoire de maîtrise ès sciences, Université de la Colombie-Britannique, Vancouver, C.-B. 131 p.
Brouwer, A., P.J.H. Reijnders et J.H. Koeman. 1989. Polychlorinated biphenyl (PCB)-contaminated fish induces vitamin A and thyroid hormone deficiency in the common seal (Phoca vitulina). Aquatic Toxicology 15: 99-106.
Brown, R.F. 1997. Pinnipeds in Oregon: status of populations and conflicts with fisheries, fish resources and human activities. p. 124-134. Dans : G. Stone, J. Goebel et S. Webster (éd). Pinniped Populations, East North Pacific: Status, Trends and Issues. 127th Annual Meeting of the American Fisheries Society, 28 août 1997, Monterey, CA.
Brown, R.F. et S.D. Reimer. 1992. Steller sea lion counts in Oregon during June and July, 1975-1991. Unpublished Report. Nongame Wildlife Program, Oregon Department of Fish and Wildlife, Newport, OR. 12 p.
Burek, K.A., F.M.D. Gulland, G. Sheffield, K.B. Beckmen, E. Keyes, T.R. Spraker, A.W. Smith, D.E. Skilling, J.F. Evermann, J.L. Stott, J.T. Saliki et A.W. Trites. 2005. Infectious disease and the decline of Steller sea lions (Eumetopias jubatus) in Alaska, USA: Insights from serologic data. Journal of Wildlife Diseases 41: 512-524.
Burek, K. A., F.M.D. Gulland, G. Sheffield, E. Keyes, T.R. Spraker, A.W. Smith, D.E. Skilling, J. Evermann, J.L. Stott et A.W. Trites. 2003. Disease agents in Steller sea lions in Alaska: a review and analysis of serology data from 1975-2000. UBC Fisheries Centre Research Reports. 11(4): 26 p.
Burkanov, V.N. 2000. Steller's sea lion population status and dynamics in Russian waters in 1989-1999. Marine Mammals of the Holarctic Regions: Materials from International Conference, Archangelsk, Russie, 21 au 23 septembre 2000. Pages 56-65 [en russe].
Burkanov, V.N. et T.R. Loughlin. 2007. Historical distribution and abundance of Steller sea lions on the Asian coast. Marine Fisheries Review. 67: 1- 62.
Calkins, D.G. 1981. Distribution and Movements of Steller Sea Lions In The Gulf of Alaska. Rapport non publié. Alaska Department of Fish and Game, Anchorage, AK. 11 pp.
Calkins, D. 1985. Steller sea lion entanglement in marine debris. p. 308-314. Dans : R.S. Shomura et H.O. Yoshida (éd.). Proceedings of the Workshop on the Fate and Impact of Marine Debris. Mémoire technique de la NOAA, NMFS-SWFC-54.
Calkins, D.G., E.F. Becker et K.W. Pitcher. 1998. Reduced body size of female Steller sea lions from a declining population in the Gulf of Alaska. Marine Mammal Science 14: 232-244.
Calkins, D., E. Becker, T.R. Spraker et T.R. Loughlin. 1994a. Impacts on Steller Sea Lions. p. 119-139. Dans : T.R. Loughlin (éd.). Marine Mammals and the Exxon Valdez. Academic Press, San Diego.
Calkins, D.G., E. Becker, T.R. Spraker et T.R. Loughlin. 1994b. Impacts on the distribution and abundance of Steller sea lions in Prince William Sound and the Gulf of Alaska. p. 119-137. Dans : T.R. Loughlin (éd.). Marine Mammals and the Exxon Valdez. Academic Press, San Diego.
Calkins, D.G. et J.A. Curatolo. 1980. Marine mammals of Lower Cook Inlet and the potential for impact from outer continental shelf oil and gas exploration, development and transport. Rapport non publié. Alaska Department of Fish and Game, Anchorage, AK. 81 p.
Calkins, D.G. et E. Goodwin. 1988. Investigation of the declining sea lion population in the Gulf of Alaska. Rapport non publié. Alaska Department of Fish and Game, Anchorage, AK. 76 p.
Calkins, D.G., D.C. McAllister, K.W. Pitcher et G.W. Pendleton. 1999. Steller sea lion status and trend in Southeast Alaska: 1979-1997. Marine Mammal Science 15: 462-477.
Calkins, D.G. et K.W. Pitcher. 1982. Population assessment, ecology and trophic relationships of Steller sea lions in the Gulf of Alaska. p. 445-546. Final Report of the Outer Continental Shelf Environmental Assessment Program. NOAA, Juneau, AK.
Calkins, D.G. et K.W. Pitcher. 1996. Steller sea lion movements, emigration and survival. pp. 34-40. Dans : K.W. Pitcher (éd.). Steller Sea Lion Recovery Investigations in Alaska, 1992-1994. ADF and G Wildlife Technical Bulletin No. 13. Alaska Department of Fish and Game, Anchorage, AK.
Colborn, T., F.S.V. Saal et A.M. Soto. 1993. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environmental Health Perspectives 101: 378-384.
Dalton, R. 2005. Is this any way to save a species? Nature 436: 14-16.
de Swart, R.L., P.S. Ross, L.J. Vedder, H.H. Timmerman, S.H. Heisterkamp, H. VanLoveren, J.G. Vos, P.J.H. Reijnders et A.D.M.E. Osterhaus. 1994. Impairment of immune function in harbor seals (Phoca vitulina) feeding on fish from polluted waters. Ambio 23: 155-159.
Deecke, V.D., P.J.B. Slater et J.K.B. Ford. 2002. Selective habituation shapes acoustic predator recognition in harbour seals. Nature. 420: 171-173.
DeLong, R.L. et G.A. Antonelis. 1991. Impacts of the 1982-1983 El Niño on the northern fur seal population at San Miguel Island, California. p. 75-83 Dans : F. Trillmich et K. Ono (éd.). Pinnipeds and El Niño: Responses to Environmental Stress. University of California Press, Berkeley, CA.
DeLong, R.L., W.G. Gilmartin et J.G. Simpson. 1973. Premature births in California sea lions: association with high organochlorine pollutant residue levels. Science 181: 1168-1170.
DeMaster, D. et S. Atkinson. (éd.). 2002. Steller sea lion decline: is it food II? Alaska Sea Grant AK-SG-02-02. 80 p.
Eberhardt, L.L. 1985. Assessing the dynamics of wild populations. Journal of Wildlife Management 49: 997-1012.
Edie, A.G. 1977. Distribution and movements of Steller sea lion cows (Eumetopias jubatus) on a pupping colony. Mémoire de maîtrise ès sciences, Université de la Colombie-Britannique, Vancouver, C.-B. 81 p.
Engelhardt, F.R. 1987. Assessment of the vulnerability of marine mammals to oil pollution. p. 101-115. Dans : J. Kuiper et W.J. Van den Brink (éd.). Fate and Effects of Oil in Marine Ecosystems. Martinus Nijoff Publishers, Boston, MA.
Engelhardt, F.R., J.R. Geraci et T.G. Smith. 1977. Uptake and clearance of petroleum hydrocarbons in the ringed seal, Phoca hispida. Journal de l’Office des recherches sur les pêcheries du Canada 34: 1143-1147.
Fiscus, C.H. et G.A. Baines. 1966. Food and feeding behaviour of Steller and California sea lions. Journal of Mammalogy 47: 195-200.
Fisher, D.H. 1981. Studies on the biology of sea lions in British Columbia. National Geographic Society Research Report 13: 215-219.
Ford, J.K.B., G.M. Ellis et J.W. Durban. 2007. Recovery potential assessment of west coast transient killer whales in British Columbia. Secrétariat canadien de consultation scientifique. Document de recherche 2007/088.
Ford, J.K.B., G.M. Ellis, L.G. Barrett-Lennard, A.B. Morton, R.S. Palm et K.C. Balcomb III. 1998. Dietary specialization in two sympatric populations of killer whales (Orcinus orca) in coastal British Columbia and adjacent waters. Revue canadienne de zoologie 76: 1456-1471.
Fowler, C.W. 1987. A review of density dependence in populations of large mammals. Current Mammalogy 1: 401-441.
Fowler, C.W. 1988. A review of seal and sea lion entanglement in marine fishing debris. pp. 16-63. Dans : D.L. Alverson et J.A. June (éd.). Proceedings of the North Pacific Rim Fishermen’s Conference on Marine Debris, Kailua-Kona, HI, 13-16 Oct 1987. Rapport non publié. National Marine Mammal Laboratory, Seattle, WA.
Fowler, C.W. et T.R. Merrell. 1986. Victims of plastic technology. Alaska Fish Game 18: 34-37.
Frid, A. et L. Dill. 2002. Human-caused disturbance stimuli as a form of predation risk. Conservation Ecology 6(1): 11.
Fritz, L.W. et E.S. Brown . 2005. Survey- and fishery-derived estimates of Pacific cod (Gadus macrocephalus) biomass: implications for strategies to reduce interactions between groundfish fisheries and Steller sea lions (Eumetopias jubatus). Fishery Bulletin 103: 501-515.
Fritz, L.W. et R.C. Ferrero. 1998. Options in Steller sea lion recovery and groundfish fishery management. Biosphere Conservation 1: 7-19.
Fritz, L.W., R.C. Ferrero et R.J. Berg. 1995. The threatened status of Steller sea lions, Eumetopias jubatus, under the Endangered Species Act: effects on Alaska groundfish fisheries management. Marine Fisheries Review 5: 14-27.
Fritz, L.W. et S. Hinckley. 2005. A critical review of the regime shift - "junk food" – nutritional stress hypothesis for the decline of the western stock of Steller sea lion. Marine Mammal Science 21(3): 476-518.
Gallucci, V., H. Nesse, I. Taylor et J. Rice. 2006. Competitive interactions among Steller sea lions, salmon sharks, and commercial fisheries. Manuscrit non publié. School of Aquatic and Fishery Sciences, Université de Washington, Seattle, WA.
Gentry, R.L. 1970. Social behaviour of the Steller sea lion. Thèse de doctorat, Université de Californie, Santa Cruz, CA. 113 p.
Gentry, R.L. et J.H. Johnson. 1981. Predation by sea lions on northern fur seal neonates. Mammalia 45: 423-430.
Gilmartin, W.G., R.L. DeLong, A.W. Smith, J.C. Sweeney, B.W. DeLappe, R.W. Risebrough, L.A. Griner, M.D.P. Dailey et D.B. Peakall. 1976. Premature parturition in the California sea lion. Journal of Wildlife Diseases 12: 104-115.
Gisiner, R.C. 1985. Male territorial and reproductive behaviour in the Steller sea lion, Eumetopias jubatus. Thèse de doctorat, Université de Californie, Santa Cruz, CA. 146 p.
Graham, G. 2004. Expert systems for marine oil spill response operations. Dans : T.W. Drosher et D.A. Fraser (éd.), 2003 GeorgiaBasin/Puget Sound Research Conference Proceedings. Puget Sound Action Team: Olympia, WA.
Grant, S.C.H. et P.S. Ross. 2002. Southern resident killer whales at risk: toxic chemicals in the British Columbia and Washington environment. Rapport technique canadien des sciences halieutiques et aquatiques 2412. 111 p.
Haebler, R. et R.B. Moeller, Jr. 1993. Pathobiology of selected marine mammal diseases. p. 217-244. Dans : J.A Couch et J.W. Fournie (éd.). Pathobiology of Marine and Estuarine Organisms. CRC Press, Boca Raton, FL.
Hall, A.J., O.I. Kalantzi et G.O. Thomas. 2003. Polybrominated diphenyl ethers (PBDEs) in grey seals during the first year of life – are they thyroid hormone endocrine disrupters? Environmental Pollution 126: 29-37.
Hancock, D. 1970. California sea lion as a regular winter visitor off the British Columbia coast. Journal of Mammalogy 51: 614.
Harding, L.E. et J.R. Englar. 1989. The Nestucca oil spill: fate and effects to May 31, 1989. Environment Canada Regional Program Report 89-01. Vancouver, C.-B.
Hare, S.R., N.J. Mantua et R.C. Francis. 1999. Inverse production regimes: Alaskan and West Coast Pacific salmon. Fisheries 24: 6-14.
Helle, E., M. Olsson et S. Jensen. 1976a. DDT and PCB levels and reproduction in ringed seal from the Bothnian Bay. Ambio 5: 188-189
Helle, E., M. Olsson et S. Jensen. 1976b. PCB levels correlated with pathological changes in seal uteri. Ambio 5: 261-263.
Henry, E. et M.O. Hammill. 2001. Impact of small boats on the haulout activity of
harbour seals (Phoca vitulina) in Metis Bay, St. Lawrence Estuary, Quebec, Canada.
Aquatic Mammals 27(2): 140-148.
Hickie, B.E., D.C.G. Muir, R.F. Addison et P. Hoekstra. 2005. Modelling the temporal trends of persistent organic pollutants in Arctic ringed seal (Phoca hispida) populations. Science of the Total Environment 351-352: 413-426.
Higgins, L. 1984. Maternal behaviour and attendance patterns of the Steller sea lion in California. Mémoire de maîtrise ès sciences, Université de Californie, Santa Cruz, CA. 37 p.
Hites, R. 2004. Polybrominated diphenyl ethers in the environment and in people: a meta-analysis of concentrations. Environmental Science and Technology 38:945-956.
Hoffman, J.I., C.W. Matson, W. Amos, T.R. Loughlin et J.W. Bickham. 2006. Deep genetic subdivision within a continuously distributed and highly vagile marine mammal, the Steller’s sea lion (Eumatopias jubatus). Molecular Ecology 15: 2821-2832.
Holmes, E.E. et A.E. York. 2003. Using age structure to detect impacts on threatened populations: a case study with Steller sea lions. Conservation Biology 17(6): 1794-1806.
Hood, W.R. et K.A. Ono. 1997. Variation in maternal attendance patterns and pup behaviour in a declining population of Steller sea lions (Eumetopias jubatus). Journal canadien de zoologie 75: 1241-1246.
Hooper, K. et T.A. McDonald. 2000. The PBDEs: an emerging environmental challenges and another reason for breast-milk monitoring programs. Environmental Health Perspectives 108: 387-392.
Hunter, A.M.J. et A.W. Trites. 2001. An annotated bibliography of scientific literature (1751-2000) pertaining to Steller sea lions (Eumetopias jubatus) in Alaska. Fisheries Centre Research Reports 9(1): 45 p.
Jameson, R.J. et K.W. Kenyon. 1977. Prey of sea lions in the Rogue River, Oregon. Journal of Mammalogy 58: 672.
Jamieson, G.S. et P.F. Olesiuk. 2001. Salmon farm - pinniped interactions in British Columbia: an analysis of predator control, its justification and alternative approaches. Document de recherche du SCCS 2001/142. 74 p.
Jepson, P.D., P.M. Bennett, R. Deaville, C.R. Allchin, J.R. Baker et R.J. Law. 2005. Relationships between polychlorinated biphenyls and health status in harbor porpoises (Phocoena phocoena) stranded in the United Kingdom. Environmental Toxicology and Chemistry 24: 238-248.
Johnson, S.R., J.J. Burns, C.I. Malme et R.A. Davis. 1989. Synthesis of information on the effects of noise and disturbance on major haulout concentrations of the Bering Sea pinnipeds. Final Report to Mineral Management Service, U.S. Department of Interior, contract no. 14-12-0001-30361. LGL Alaska Research Associates, Anchorage, AK. 284 p.
Kajimura, H. et T.R. Loughlin. 1988. Marine mammals in the oceanic food web of the eastern subarctic Pacific. Bulletin of the Ocean Research Institue, Université de Tokyo 26: 187-223.
Kannan, K., J. Koistinen, K. Beckmen, T. Evans, J.F. Gorzelany, K.J. Hansen, P.D. Jones, E. Helle, M. Nyman et J.P. Giesy. 2001. Accumulation of perfluorooctane sulfonate in marine mammals. Environmental Science and Technology 35: 1593-1598.
Kastelein, R.A., N. Vaughan et P.R. Wiepkema. 1990. The food consumption of Steller sea lions (Eumetopias jubatus). Aquatic Mammals 15: 137-144.
Kenyon, K.W. 1962. History of the Steller sea lion at the Pribilof Islands, Alaska. Journal of Mammalogy 43: 68-75.
Kenyon, K.W. et D.W. Rice. 1961. Abundance and distribution of the Steller sea lion. Journal of Mammalogy 42: 223-234.
Kucey, L. 2005. Human disturbance and the hauling out behaviour of Steller sea lions (Eumetopias jubatus). Mémoire de maîtrise ès sciences, Université de la Colombie-Britannique, Vancouver, C.-B. 67 p.
Lambourn, D.M., S.J. Jeffries, M.M. Lance, P.F. Olesiuk, S.A. Raverty et A.W. Trites. 2006. Blood analysis on free ranging Steller sea lions (Eumetopias jubatus). International Association for Aquatic Animal Medicine Conference, 6-10 May 2006. Nassau, Bahamas.
Lee, J.S., S. Tanabe, H. Umino, R. Tatsukawa, T.R. Loughlin et D.C. Calkins. 1996. Persistent organochlorines in Steller sea lion (Eumetopias jubatus) from the bulk of Alaska and the Bering Sea, 1976-1981. Marine Pollution Bulletin 32: 535-544.
Lewis, J.P. 1987. An evaluation of a census-related disturbance of Steller sea lions. Mémoire de maîtrise ès sciences, Université de l’Alaska, Fairbanks, AK. 93 p.
Lindstrom, G., H. Wingfors, M. Dam et B. von Bavel. 1999. Identification of 19 polybrominated diphenyl ethers (PBDEs) in long-finned pilot whale (Globicephala melas) from the Atlantic. Archives of Environmental Contamination and Toxicology 36: 355-363.
Livingston, P.A. 1991. Groundfish food habits and predation on commercially important prey species in the eastern Bering Sea from 1984-1986. Mémoire technique de la NOAA, NMFS-NWFC-207.
Loughlin, T.R. 1997. Using the phylogeographic method to identify Steller sea lion stocks. pp. 159-171. Dans : A. Dizon, S.J. Chivers et W.F. Perrin (éd.). Molecular genetics of marine mammals. Special Publication #3 of the Society for Marine Mammalogy.
Loughlin, T.R. 1998. The Steller sea lion: A declining species. Biosphere Conservation 1: 91-98.
Loughlin, T.R., B.E. Ballachey et B.A. Wright. 1996. Overview of studies to determine injury caused by the Exxon Valdez oil spill to marine mammals. American Fisheries Society Symposium 18: 798-808.
Loughlin, T.R., L. Consiglieri, R.L. DeLong et A.T. Actor. 1983. Incidental catch of marine mammals by foreign fishing vessels, 1978-1981. Marine Fisheries Review 45: 44-49.
Loughlin, T.R. et R.L. DeLong. 1983. Incidental catch of sea lions during the 1982 and 1983 walleye pollock joint venture fishery in Shelikof Strait, Alaska. NWAFC Processed Report. 83-15. 37 p.
Loughlin, T.R., P.J. Gearin, R.L. DeLong et R.L. Merrick. 1986. Assessment of net entanglement on northern sea lions in the Aleutian Islands, 25 June-15 July 1985. NWAFC Processed Rep. 86-02. National Marine Mammal Laboratory, Seattle WA. 50 p.
Loughlin, T.R. et R. Nelson, Jr. 1986. Incidental mortality of northern sea lions in Shelikof Strait, Alaska. Marine Mammal Science 2: 14-33.
Loughlin, T.R., M.A. Perez et R.L. Merrick. 1987. Eumetopias jubatus. Mammalian Species 283: 1-7.
Loughlin, T.R., A.S. Perlov, J.D. Baker, S.A. Blokhin et A.G. Makhnyr. 1998. Diving behaviour of adult female Steller sea lions in the Kuril Islands, Russia. Biosphere Conservation 1: 21-31.
Loughlin, T.R., A.S. Perlov et V.A. Vladimirov. 1992. Range-wide survey and estimation of total number of Steller sea lions in 1989. Marine Mammal Science 8: 220-239.
Loughlin, T.R., D.J. Rugh et C.H. Fiscus. 1984. Northern sea lion distribution and abundance: 1956-1980. Journal of Wildlife Management 48: 729-740.
Loughlin, T.R., J T. Sterling, R.L. Merrick, J.L. Sease et A.E. York. 2003. Diving behaviour of immature Steller sea lions (Eumetopias jubatus). Fishery Bulletin 101: 566-582.
Loughlin, T.R. et A.E. York. 2000. An accounting of the sources of Steller sea lion, Eumetopias jubatus, mortality. Marine Fisheries Review 62: 40-45.
Lowe, S.A. et L.W. Fritz. 1997. Atka mackerel. Stock assessment and fishery evaluation report for the groundfish resources of the Bering Sea/Aleutian Islands regions for 1998. North Pacific Fisheries Management Council, Anchorage, AK.
Lowry, M. et O. Maravilla-Chavez. 2005. Recent abundance of California sea lions in western Baja California, Mexico and the United States. Proceedings of the 6th California Islands Symposium, Ventura, CA, 1-3 Dec 2003. National Park Service Technical Publication CHIS-05-01, Institute for Wildlife Studies, Arcata, CA.
Maniscalco, J.M., C.O. Matkin, D. Maldini, D.G. Calkins et S. Atkinson. 2007. Assessing killer whale predation on Steller sea lions from field observations in Kenai Fjords, Alaska. Marine Mammal Science 23: 306-321.
Maniscalco, J.M., K. Wynne, K.W. Pitcher, M.B. Hanson, S.R. Melin et S. Atkinson. 2004. The occurrence of California sea lions (Zalophus californianus) in Alaska. Aquatic Mammals 30: 427-433.
Martin, J.H., P.D. Elliot, V.C. Anderlini, D. Girvin, S.A. Jacobs, R.W. Risebrough, R.L. DeLong et W.G. Gilmartin. 1976. Mercury-selenium-bromine imbalance in premature parturient California sea lions. Marine Biology 35: 91-104.
Mate, B.R. 1975. Annual migration of the Steller sea lion and California sea lion along the coast of Oregon. Journal du Conseil international pour l’exploration de la mer 169: 455-461.
Mate, B.R. 1985. Incidents of marine mammal encounters with debris and active fishing gear. p. 453-457. Dans : R.S. Shomura et H. O. Yoshida (éd.). Proceedings of the Workshop on the Fate and Impact of Marine Debris. Mémoire technique de la NOAA, NMFS-SWFC-54.
Mathisen, O.A., R.T. Baade et R.J. Lopp. 1962. Breeding habits, growth, and stomach contents of the Steller sea lion in Alaska. Journal of Mammalogy 43: 464-477.
Matkin, C.O., L.G. Barrett-Lennard, H. Yurk, D. Ellifrit et A.W. Trites. 2007. Ecotypic variation and predatory behaviour among killer whales (Orcinus orca) off the eastern Aleutian Islands, Alaska. Fishery Bulletin 105: 74-87.
McAlister, W.B. et M.A. Perez. 1976. Ecosystem dynamics, birds and marine mammals. Part I: Preliminary estimates of pinniped-finfish relationships in the Bering Sea. Final report for Environmental Assessment of the Alaskan Continental Shelf. Alaska Fisheries Science Center. Seattle, WA. p. 219-248.
McFarlane, G.A., J.R. King et R.J. Beamish. 2000. Have there been recent changes in climate? Ask the fish. Progress in Oceanography 47: 147-169.
Melin, S.R. et R.L. DeLong. 1994. Population monitoring of northern fur seals on San Miguel Island, California. p. 137-141. Dans : E.H. Sinclair (éd.). Fur Seal Investigations, 1992. Mémoire technique de la NOAA, NMFS-AFSC-45. 190 p.
Melin, S.R. et R.L. DeLong. 2000. Population monitoring studies of northern fur seals at San Miguel Island, California. p. 41-51. Dans : B.W. Robson (éd.). Fur Seal Investigations, 1998. Mémoire technique de la NOAA, NMFS-AFSC-113. 101 p.
Melin, S.R., R.L. DeLong et J.R. Thomason. 1996. Population monitoring studies of northern fur seals at San Miguel Island, California. p. 87-102. Dans : E.H. Sinclair (éd.). Fur Seal Investigations, 1994. Mémoire technique de la NOAA, NMFS-AFSC-69. 144 p.
Merrick, R.L. 1987. Behavioural and demographic characteristics of northern sea lion rookeries. Mémoire de maîtrise ès sciences, Université d’État de l’Oregon, Corvallis, OR. 124 p.
Merrick, R.L. 1997. Current and historical roles of apex predators in the Bering Sea ecosystem. Journal of Northwest Atlantic Fishery Science: 343-356.
Merrick, R.L., M.K. Chumbley et G.V. Byrd. 1997. Diet diversity of Steller sea lions (Eumetopias jubatus) and their population decline in Alaska: a potential relationship. Journal canadien des sciences halieutiques et aquatiques 54: 1342-1348.
Merrick, R.L. et T.R. Loughlin. 1997. Foraging behaviour of adult female and young-of-year Steller sea lions in Alaskan waters. Revue canadienne de zoologie 75: 776-786.
Milette, L.L. 1999. Behaviour of lactating Steller sea lions (Eumetopias jubatus) during the breeding season: A comparison between a declining and stable population in Alaska. Mémoire de maîtrise ès sciences, Université de la Colombie-Britannique, Vancouver, C.-B. 57 p.
Milette, L.L. et A.W. Trites. 2003. Maternal attendance patterns of lactating Steller sea lions (Eumetopias jubatus)from a stable and a declining population in Alaska. Revue canadienne de zoologie 81: 340-348.
Morton, A.B. 1990. A quantitative comparison of the behaviour of resident and transient forms of the killer whale off the central British Columbia coast. Rapport de la Commission baleinière internationale, Numéro spécial 12: 245-248.
Mos, L., B. Morsey, S.J. Jeffries, M.B. Yunker, S. Raverty, S. De Guise et P.S. Ross. 2006. Chemical and biological pollution contribute to the immunological profiles of free-ranging harbor seals. Environmental Toxicology and Chemistry 25: 3110-3117.
Mos, L., M. Tabuchi, N. Dangerfield, S.J. Jeffries, B.F. Koop et P.S. Ross. 2007. Contaminant-associated disruption of vitamin A and its receptor (RARa) in free-ranging harbour seals (Phoca vitulina). Aquatic Toxicology 81: 319-328.
MPO. 2007. La situation des pêches en Colombie-Britannique, Tendances des prises commerciales en C.-B. (1996-2005). Disponible à l’adresse : http://www.pac.dfo-mpo.gc.ca/ops/fm/reports/fisheries05/stats_f.htm [consulté le 28 octobre 2007].
MPO. 2007b. Programme de rétablissement de la loutre de mer (Enhydra lutris) au Canada [version finale]. Série de Programmes de rétablissement de la Loi sur les espèces en péril. Pêches et Océans Canada, Vancouver. vi + 46 p. Disponible à l’adresse : http://www.registrelep.gc.ca
MPO. 2007c. Programme de rétablissement de l’épaulard migrateur (Orcinus orca) au Canada [version finale]. Série de Programmes de rétablissement de la Loi sur les espèces en péril. Pêches et Océans Canada, Vancouver. vi + 46 p.
MPO. 2008. Évaluation des populations d’otarie de Steller (Eumetopias jubatus). Secr. can. de consult. sci. du MPO. Avis sci. 2008/047.
MPO. 2008b. Plan de gestion de la baleine grise de l’est du Pacifique (Eschrichtius robustus) dans les eaux canadiennes du Pacifique [Proposition]. Série de Plans de gestion de la Loi sur les espèces en péril. Pêches et Océans Canada, Nanaimo. Disponible à l’adresse : http://www-comm.pac.dfo-mpo.gc.ca/pages/consultations/consult_f.htm.
MPO. 2008c. Programme de rétablissement des épaulards résidents (Orcinus orca) du nord et du sud au Canada [version finale]. Série de Programmes de rétablissement de la Loi sur les espèces en péril. Pêches et Océans Canada, Ottawa. 76 + ix p. Disponible à l’adresse : http://www.registrelep.gc.ca.
MPO. 2009. Plan de gestion de l’épaulard du large (Orcinus orca) au Canada [version finale]. Série de Plans de gestion de la Loi sur les espèces en péril. Pêches et Océans Canada, Nanaimo. ix + 51 p. Disponible à l’adresse : http://www.registrelep.gc.ca.
MPO. 2009b. Plan de gestion du marsouin commun du Pacifique (Phocoena phocoena) au Canada [version finale].Série de Plans de gestion de la Loi sur les espèces en péril. Pêches et Océans Canada, Nanaimo. Disponible à l’adresse : http://www.registrelep.gc.ca.
Newcombe, C.F. et W.A. Newcombe. 1914. Sea-lions on the coast of British Columbia, Report of the Commission of Fisheries for 1913. 131-139 p.
NMFS (National Marine Fisheries Service). 1992. Recovery Plan for the Steller sea lion (Eumetopias jubatus). Prepared by the Steller Sea Lion Recovery Team for the Office of Protected Resources, National Marine Fisheries Service, Silver Spring, MD. 92 p.
NMFS (National Marine Fisheries Service). 2001. Endangered Species Act, Section 7 Consultation Biological Opinion and Incidental Take Statement on the authorization of the Bering Sea/Aleutian Islands and Gulf of Alaska Groundfish Fishery Management Plan Amendments 61 and 70. NMFS Alaska Region, Protected Resources Division, Juneau, AK. 206 p.
NMFS (National Marine Fisheries Service). 2008. Steller sea lion recovery plan, eastern and western distinct population segments (Eumetopias jubatus). Revision. Office of Protected Resources, National Marine Fisheries Service, Silver Spring, MD. 325 p.
O’Corry-Crowe, G., T. Gelatt, K. Pitcher et B. Taylor. 2005. Crossing significant boundaries: evidence of mixed-stock origins of new Steller sea lion, Eumetopias jubatus, rookeries in southeast Alaska. 6th Biennial Conference on the Biology of Marine Mammals, 12-16 Dec 2005, San Diego, CA.
O’Corry-Crowe, G., B.L. Taylor, T. Gelatt, T.R. Loughlin, J. Bickman, M. Basterrretche, K.W. Pitcher et D.P. DeMaster. 2007 Demographic independence along ecosystem boundaries in Steller sea lions revealed by mtDNA analysis: implications for management of an endangered species. Revue canadienne de zoologie 84: 1796-1809.
O’Daniel, D. et J.C. Schneeweis. 1992. Steller sea lion, Eumetopias jubatus, predation on Glaucous-winged gulls, Larus glaucescens. Canadian Field-Naturalist 106: 268.
Olesiuk, P.F. 1999. An assessment of the status of harbour seals (Phoca vitulina) in British Columbia. Document de recherche du Secrétariat canadien pour l’évaluation des stocks. 1999/33. 71 p.
Olesiuk, P.F. 2008. Abondance de l’otarie de Steller (Eumetopias jubatus) en Colombie-Britannique. Document de recherche du secrétariat canadien de consultation scientifique. 2008/063.
Olesiuk, P.F., D.G. Calkins, K.W. Pitcher, W.L. Perryman, C. Stinchcomb et M. Lynn. 2008. Évaluation des dénombrements de petits de l’otarie de Steller (Eumetopias jubatus) à partir d’images 35mm prises à angle oblique. Document de recherche du Secrétariat canadien de consultation scientifique. 2008/064.
Olesiuk, P.F. et M.A. Bigg. 1988. Seals and sea lions in British Columbia. Department of Fisheries and Oceans Special Publication. Station biologique du Pacifique, Nanaimo, C.-B. 12 p.
Olesiuk, P.F., M.A. Bigg et G.M. Ellis. 1990. Recent trends in the abundance of harbour seals, Phoca vitulina, in British Columbia. Journal canadien des sciences halieutiques et aquatiques 47(5): 992-1003.
Orr, R. T. et T.C. Poulter. 1967. Some observations on reproduction, growth, and social behaviour in the Steller sea lion. Proceedings of the California Academy of Science 35: 193-226.
Pascual, M.A. et M.D. Adkinson. 1994. The decline of the Steller sea lion in the northeast Pacific: demography, harvest or environment? Ecological Applications 4: 393-403.
Pauly, D., V. Christensen, J. Dalsgaard, R. Froese et F. Torres Jr. 1998. Fishing down marine food webs. Science 279: 860-863.
Pearson, J.P. et J.P. Verts. 1970. Abundance and distribution of harbor seals and northern sea lions in Oregon. Murrelet 51: 1-5.
Perez, M.A. 1994. Calorimetry measurements of energy value of some Alaskan fishes and squids. Mémoire technique de la NOAA, NMFS-AFSC-32. 32 p.
Pike, G.C. 1958. Food of the northern sea lion. Conseil de recherches sur les pêcheries du Canada, Rapport d’étape de la Station biologique de la côte du Pacifique. 112, 18-20 p.
Pike, G.C. et B.E. Maxwell. 1958. The abundance and distribution of the northern sea lion (Eumetopias jubata) on the coast of British Columbia. Journal de l’Office des recherches sur les pêcheries du Canada 15: 5-17.
Pitcher, K.W. 1981. Prey of the Steller sea lion, Eumetopias jubatus, in the Gulf of Alaska. Fishery Bulletin 79: 467-472.
Pitcher, K.W., D.G. Calkins et G. Pendleton. 1998. Reproductive performance of female Steller sea lions: an energetics-based reproductive strategy? Revue canadienne de zoologie 76: 2075-2083.
Pitcher, K.W. et F.H. Fay. 1982. Feeding by Steller sea lions on harbor seals. Murrelet 63: 70-71.
Pitcher, K.W., P.F. Olesiuk, R.F. Brown, M. Lowry, J. Sease, W. Perryman, C. Stinchcomb et L. Lowry. 2003. Status and trend of the eastern population of Steller sea lions. Marine Sciences in the Northeast Pacific Symposium, 13-17 Jan 2003, Anchorage, AK.
Pitcher, K.W., P.F. Olesiuk, R.F. Brown, M.S. Lowry, S.J. Jeffries, J.L. Sease, W.L. Perryman, C.E. Stinchcomb et L.F. Lowry. 2007. Abundance and distribution of the eastern North Pacific Steller sea lion (Eumetopias jubatus) population. Fishery Bulletin 105: 102-115.
Raum-Suryan, K. et K.W. Pitcher. 2000. Trip report: brand resights of Steller sea lions within Southeast Alaska and northern British Columbia from 19 June - 10 July, 2000. Rapport non publié. Alaska Department of Fish and Game, Anchorage, AK.
Raum-Suryan, K.L., K.W. Pitcher, D.G. Calkins, J.L. Sease et T.R. Loughlin. 2002. Dispersal, rookery fidelity, and metapopulation structure of Steller sea lions (Eumetopias jubatus) in an increasing and a decreasing population in Alaska. Marine Mammal Science 18: 746-764.
Rayne, S., M.G. Ikonomou, P.S. Ross, G.M. Ellis et L.G. Barrett-Lennard. 2004. PBDEs, PBBs, and PCNs in three communities of free-ranging killer whales (Orcinus orca) from the northeastern Pacific Ocean. Environmental Science and Technology 38: 4293-4299.
Reijnders, P.J.H. 1986. Reproductive failure in common seals feeding on fish from polluted coastal waters. Nature 324: 456-457.
Rice, D.W. 1998. Marine mammals of the world: systematics and distribution. Special Publication of the Society of Marine Mammalogy. 231 p.
Roffe, T.J. et B.R. Mate. 1984. Abundances and feeding habits of pinnipeds in the Rogue River, Oregon. Journal of Wildlife Management 48: 1261-1274.
Rosen, D.A.S. et A.W. Trites. 1999. Metabolic effects of low-energy diet on Steller sea lions, Eumetopias jubatus. Physiological and Biochemical Zoology 72: 723-731.
Rosen, D.A.S. et A.W. Trites. 2000a. Assessing the role of nutritional stress in the decline of wild populations: a Steller case of scientific sleuthing. p. 182-186. Dans : C.L.K. Baer (éd.). Proceedings of the Third Comparative Nutrition Society Symposium, Pacific Grove, CA, 4-9 August 2000.
Rosen, D.A.S. et A.W. Trites. 2000b. Digestive efficiency and dry matter digestibility of Steller sea lions fed herring, pollock, squid, and salmon. Revue canadienne de zoologie 78: 234-239.
Rosen, D.A.S. et A.W. Trites. 2000c. Pollock and the decline of Steller sea lions: testing the junk-food hypothesis. Revue canadienne de zoologie 78: 1243-1258.
Ross, P.S. 2000. Marine mammals as sentinels in ecological risk assessment. Human and Ecological Risk Assessment 6: 29-46
Ross, P.S. 2002. The role of immunotoxic environmental contaminants in facilitating the emergence of infectious diseases in marine mammals. Human and Ecological Risk Assessment 8(2): 277-292.
Ross, P.S. 2006. Fireproof killer whales (Orcinus orca): Flame retardant chemicals and the conservation imperative in the charismatic icon of British Columbia, Canada. Journal canadien des sciences halieutiques et aquatiques 63: 224-234.
Ross, P.S., R.L. de Swart, P.J.H. Reijnders, H.V. Loveren, J.G. Vos et A.D.M.E. Osterhaus. 1995. Contaminant-related suppression of delayed-type hypersensitivity and antibody responses in harbour seals fed herring from the Baltic Sea. Environmental Health Perspectives 103: 162-167.
Ross, P.S., R.L. de Swart, H.H. Timmerman, H. van Loveren, J.G. Vos et A.D.M.E. Osterhaus. 1996. Immunotoxicological studies in the harbour seal (Phoca vitulina). p. 579-584. Dans : J.S. Stolen, T.C. Fletcher, C.J. Bayne, C.J. Secombes, J.T. Zelikoff, L.E. Twerdok et D.P. Anderson (éd.). Modulators of Immune Responses: the Evolutionary Trail. SOS Publications, Fair Haven, NJ.
Ross, P.S., R.L. De Swart, H. Van der Vliet, L. Willemsen, A. De Klerk, G. Van Amerongen, J. Groen, A. Brouwer, I. Schipholt, D.C. Morse, H. Van Loveren, A.D.M.E. Osterhaus et J.G. Vos. 1997. Impaired cellular immune response in rats exposed perinatally to Baltic Sea herring oil or 2,3,7,8-TCDD. Archives of Toxicology 17: 563-574
Rowley, J. 1929. Life history of sea lions on the California coast. Journal of Mammalogy 10: 1-36.
Sandegren, F.E. 1970. Breeding and maternal behaviour of the Steller sea lion (Eumetopias jubatus) in Alaska. Mémoire de maîtrise ès sciences, Université de l’Alaska, Fairbanks, AK 138 p.
Sandegren, F.E. 1976. Courtship display, agonistic behaviour and social dynamics in the Steller sea lion. Behaviour 57: 136-158.
Scheffer, V.B. 1964. Hair patterns in seals (Pinnipedia). Journal of Morphology 115: 291-303.
Schusterman, R.J. 1981. Steller sea lion, Eumetopias jubatus (Schreber, 1776). p. 119-141. Dans : S.H. Ridgway et R.J. Harrison (éd.). Handbook of Marine Mammals. Vol. 1. Walrus, Sea Lions, Fur Seals and Sea Otters. Academic Press, New York, NY.
Scordino, J. 2006. Steller sea lions (Eumetopias jubatus) of Oregon and northern California: seasonal haulout abundance patterns, movements of marked juveniles, and effects of hot-branding on apparent survival of pups at Rogue Reef. Mémoire de maîtrise ès sciences, Université de l’Oregon, Corvallis, OR. 93 p.
Sease, J.L., R.F. Brown, V.N. Burkanov, D.G. Calkins, P.F. Olesiuk et A.E. York. 1999. Range-wide survey of Steller sea lions in 1994. Rapport non publié. National Marine Mammal Laboratory, Seattle, WA.
Sease, J.L. et C. Stinchcomb. 2003. 2002 Surveys of Steller Sea Lions in Alaska. Marine Sciences in the Northeast Pacific Symposium, 13-17 Jan 2003, Anchorage, AK.
Sheffield, G. et R. Zarnke. 1997. Summaries of serologic data collected from Steller sea lions in the Bering Sea and Gulf of Alaska, 1978-1996. p. 74-84. Dans : K.W. Pitcher (éd.). Steller Sea Lion Recovery Investigations in Alaska, 1995-1996. NOAA Contract Report NA57FX0256. Alaska Department of Fish and Game, Division of Wildlife Conservation, Anchorage, AK.
Shima, M., A.B. Hollowed et G.R. VanBlaricom. 2000. Response of pinniped populations to directed harvest, climate variability, and commercial fishery activity: a comparative analysis. Reviews in Fisheries Science 8: 89-124.
Sigler, M.F., L.B. Hulbert, C.R. Lunsford, N.H. Thompson, K. Burek, G. O’Corry- Crowe et A.C. Hirons. 2006. Diet of Pacific sleeper shark, a potential Steller sea lion predator, in the northeast Pacific Ocean. Journal of Fish Biology 69: 392-405.
Sinclair, E., T.R. Loughlin et W. Pearcy. 1994. Prey selection by northern fur seals (Callorhinus ursinus) in the eastern Bering Sea. Fishery Bulletin 92: 144-156.
Sinclair, E.H., G.A. Antonelis, B.W. Robson, R.R. Ream et T.R. Loughlin. 1996. Northern fur seal, Callorhinus ursinus, predation on juvenile walleye pollock, Theragra chalcogramma. p. 167-178. Dans : R.D. Brodeur, P.A. Livingston, T.R. Loughlin et A.B. Hollowed (éd.). Ecology of Juvenile Walleye Pollock, Theragra chalcogramma. NOAA Technical Report NMFS 126.
Sinclair, E.H. et T.K. Zeppelin. 2002. Seasonal and spatial differences in diet in the western stock of Steller sea lions (Eumetopias jubatus). Journal of Mammalogy 83: 973-990.
Smith, T.G. et J.R. Geraci. 1975. The effect of contact and ingestion of crude oil on ringed seals in the Beaufort Sea. Beaufort Sea Technical Report No. 5. 66 p.
Song, L., A. Seeger et J. Santos-Such. 2005. On membrane motor activity and chloride flux in the outer hair cell: lessons learned from the environmental toxin tributyltin. Biophysical Journal 88 (3): 2350-2362.
Spalding, D.J. 1964. Comparative feeding habits of the fur seal, sea lion and harbour seal on the British Columbia coast. Journal de l’Office des recherches sur les pêcheries du Canada 146: 1-47.
Springer, A.M. 1998. Is it all climate change? Why marine bird and mammal populations fluctuate in the North Pacific. p. 109-119. Dans : G. Holloway, P. Muller et D. Henderson (éd.). Biotic Impacts of Extratropical Climate Variability in the Pacific. Proceedings of the 'Aha Huliko'a Hawaiian Winter Workshop. 26-29 Jan 1998. Université d’Hawaii, Honolulu, HI.
St. Aubin, D.J. 1990. Physiologic and toxic effects on pinnipeds. p. 103-127. Dans : J.R. Geraci et D.J. St. Aubin (éd.). Sea Mammals and Oil: Confronting the Risks. Academic Press, San Diego, CA.
Stewart, B.S. et P.K. Yochem. 1987. Entanglement of pinnipeds in synthetic debris and fishing net and line fragments at San Nicolas and San Miguel Islands, California, 1978-1986. Marine Pollution Bulletin 18: 336-339.
Stroud, R.K. 1978. Causes of death and pathological findings in marine mammals along the Oregon coast. Mémoire de maîtrise ès sciences, Université d’État de l’Oregon, Corvalis, OR. 65 p.
Swain, U.G. 1996. Foraging behaviour of female Steller sea lions in Southeast Alaska and the eastern Gulf of Alaska. p. 135-166. Steller sea lion recovery investigations in Alaska, 1992-1994. Wildlife Technical Bulletin 13, May 1996. Alaska Department of Fish and Game, Anchorage, AK.
Swain, U.G. 1999. Steller sea lion foraging studies in Southeast Alaska during 1998. Steller Sea Lion Research Peer Review Feeding Ecology Workshop. 11-12 Feb 1999. Seattle, WA. 40 p.
Swain, U. et D.G. Calkins. 1997. Foraging behaviour of juvenile Steller sea lions in the northeastern Gulf of Alaska: diving and foraging trip duration. p. 92-106. Steller sea lion recovery investigations in Alaska, 1995-1996. Rapport non publié. Alaska Department of Fish and Game, Anchorage, AK.
Szaniszlo, W. 2005. California sea lion (Zalophus californianus) and Steller sea lion (Eumetopias jubatus) interactions with vessels in Pacific Rim National Park Reserve: implications for marine mammal viewing management. Mémoire de maîtrise ès sciences. Université de Victoria.
Tabuchi, M., N. Veldhoen, N. Dangerfield, S.J. Jeffries, C.C. Helbing et P.S. Ross. 2006. PCB-related alteration of thyroid hormones and thyroid hormone receptor gene expression in free-ranging harbor seals (Phoca vitulina). Environmental Health Perspectives 114: 1024-1031.
Tamura, T. et S. Ohsumi. 2000. Regional assessments of prey consumption by marine cetaceans in the world. Commission baleinière internationale, Scientific Committee Document SC/52/E6. 42 p.
Thorsteinson, F.V. et C.J. Lensink. 1962. Biological observations of Steller sea lions taken during an experimental harvest. Journal of Wildlife Management 26: 353-359.
Trenberth, K.E. 1990. Recent observed interdecadal climate changes in the northern hemisphere. Bulletin of the American Meteorological Society 71: 988-993.
Trites, A.W. 1997. The role of pinnipeds in the ecosystem. p. 31-39. Dans : G. Stone, J. Goebel et S. Webster (éd.). Pinniped populations, eastern north Pacific: status, trends and issues, New England Aquarium, Conservation Department, Boston, MA.
Trites, A.W., V. Christensen et D. Pauly. 2006. Effects of fisheries on ecosystems: just another top predator? p. 11-27. Dans : I.L. Boyd, K. Camphuysen et S. Wanless (éd.). Top Predators in Marine Ecosystems: Their Role in Monitoring and Management. Cambridge University Press, Cambridge.
Trites, A.W. et C.P. Donnelly. 2003. The decline of Steller sea lions in Alaska: a review of the nutritional stress hypothesis. Mammal Review 33: 3-28.
Trites, A.W. et P.A. Larkin. 1996. Changes in the abundance of Steller sea lions (Eumetopias jubatus) in Alaska from 1956 to 1992: How many were there? Aquatic Mammals 22: 153-166.
Trites, A.W., P.A. Livingston, S. Mackinson, M.C. Vasconcellos, A.M. Springer et D. Pauly. 1999. Ecosystem change and the decline of marine mammals in the eastern Bering Sea: testing the ecosystem shift and commercial whaling hypotheses. UBC Fisheries Centre Research Reports. 7(1): 106 p.
Trites, A.W., A.J. Miller, H.D.G. Maschner, M.A. Alexander, S.J. Bograd, J.A. Calder, A. Capotondi, K.O. Coyle, E.D. Lorenzo, B.P. Finney, E.J. Gregr, C.E. Grosch, S.R. Hare, G.L. Hunt, J. Jahncke, N.B. Kachel, H.-J. Kim, C. Ladd, N.J. Mantua, C. Marzban, W. Maslowski, R. Mendelssohn, D.J. Neilson, S.R. Okkonen, J.E. Overland, K.L. Reedy-Maschner, T.C. Royer, F.B. Schwing, J.X.L. Wang et A.J. Winship. 2007. Bottom-up forcing and the decline of Steller sea lions in Alaska: assessing the ocean climate hypothesis. Fisheries Oceanography 16: 46-67.
Van de Vijver, K.I., P.T. Hoff, K. Das, W. Van Dongen, E.L Esmans, T. Jauniaux, J. Bouquegenau, R. Blust et W. de Coen. 2003. Perfluorinated chemicals infiltrate ocean waters: link between exposure levels and stable isotope ratios in marine mammals. Environmental Science and Technology 37: 5545-5550.
Wada, K. 1998. Steller sea lions: present status of studies of migratory ecology, and conflict between fisheries and conservation in Japan. Biosphere Conservation 1: 1-6.
Wade, P., V.N. Burkanov, M.E. Dahlheim, N.A. Friday, L.W. Fritz, T.R. Loughlin, S.A. Mizroch, M.M. Muto, D.W. Rice, L.G. Barrett-Lennard, N.A. Black, A.M. Burdin, J. Calambokidis, S. Cerchio, J.K.B. Ford, J.K. Jacobsen, C.O. Matkin, D.R. Matkin, A.V. Mehta, R.J. Small, J.M. Straley, S.M. McCluskey et G.R. VanBlaricom. 2007. Killer whales and marine mammal trends in the North Pacific – A re-examination of evidence for the sequential megafauna collapse and the prey-switching hypothesis. Marine Mammal Science 23(4): 766-802.
Wespestad, V.G., L.W. Fritz, W.J. Ingraham et B.A. Megrey. 2000. On relationships between cannibalism, climate variability, physical transport, and recruitment success of Bering Sea walleye pollock (Theragra chalcogramma). ICES Journal of Marine Science 57: 272-278.
Wilke, F. et K.W. Kenyon. 1952. Notes on the food of fur seal, sea-lion, and harbor porpoise. Journal of Wildlife Management 16: 396-397.
Winship, A.J. et A.W. Trites. 2003. Prey consumption of Steller sea lions (Eumetopias jubatus) off Alaska: How much prey do they require? Fishery Bulletin 101: 147-167.
Winship, A.J., A.W. Trites et D.G. Calkins. 2001. Growth in body size of the Steller sea lion (Eumetopias jubatus). Journal of Mammalogy 82: 500-519.
Winship, A.J., A.W. Trites et D.A.S. Rosen. 2002. A bioenergetic model for estimating the food requirements of Steller sea lions (Eumetopias jubatus) in Alaska, USA. Marine Ecology Progress Series 229: 291-312.
Withrow, D.E. 1982. Using aerial surveys, ground truth methodology, and haul out behaviour to census Steller sea lions, Eumetopias jubatus. Mémoire de maîtrise ès sciences, Université de Washington, Seattle, WA. 102 p.
York, A.E. 1994. The population dynamics of northern sea lions, 1975-1985. Marine Mammal Science 10: 38-51.
York, A.E., R.L. Merrick et T.R. Loughlin. 1996. An analysis of the Steller sea lion metapopulation in Alaska. p. 259-292. Dans : D.R. McCullough (éd.). Metapopulations and Wildlife Conservation. Island Press, Washington, DC.
Tableau 4. Détails sur les termes utilisés pour l’évaluation des menaces pesant sur la population d’otaries de Steller de l’est du Pacifique.
| TERMES | CLASSIFICATION | DÉFINITIONS |
| Incertitude | Faible | L’effet de la menace présente un lien causal avec la diminution de la viabilité de la population et est susceptible d’entraîner l’échec de l’atteinte des objectifs du plan de gestion. |
| Moyenne | L’effet de la menace est corrélé avec la diminution de la viabilité de la population et a une incidence négative sur les objectifs du plan de gestion. | |
| Élevée | L’effet négatif de la menace sur la viabilité de la population ou sur l’atteinte des objectifs du plan de gestion est présumé ou plausible. | |
| Gravité | Négligeable | La menace n’a pas d’effet observable sur la population. |
| Faible | Les effets de la menace sont sublétaux et entraîneront probablement des changements de comportement à court terme. | |
| Modérée | Les effets de la menace entraînent des changements comportementaux ou physiologiques chroniques (p. ex. possibilité de déplacement à long terme hors de l’habitat). | |
| Élevée | Les effets de la menace sont létaux. | |
| Inconnue | L’information disponible est insuffisante pour que l’on puisse établir la mesure dans laquelle la menace peut affecter la viabilité de la population. | |
| Potentiel d’atténuation |
Faible | La mise en œuvre de mesures pour atténuer ou éviter les impacts sur la viabilité de la population est impossible ou sera vraisemblablement un échec. |
| Modéré | La mise en œuvre de mesures pour atténuer ou éviter les impacts sur la viabilité de la population est faisable et susceptible de donner certains résultats. | |
| Élevé | La mise en œuvre de mesures pour atténuer ou éviter les impacts sur la viabilité de la population est en cours et il devrait être très facile d’appliquer, à l’avenir, des mesures qui seront vraisemblablement très fructueuses. | |
| Inconnu | L’information disponible ne nous permet pas d’établir s’il est possible d’atténuer les effets de la menace. |
Tableau 5. Polluants organiques persistants qui peuvent poser un risque pour l’otarie de Steller. Ce tableau provient de la version finale du programme de rétablissement des épaulards résidents au Canada (MPO, 2008).
Polluant |
Utilisation/source | Persistant | Bioaccumulatif | Risques |
| DDT Dichlorodi-phényle trichloroéthane |
Pesticide utilisé dans certains pays, banni en Amérique du Nord, persiste dans les eaux de ruissellement 30 ans après l’interdiction d’usage, pénètre dans l’atmosphère à partir des régions où il est encore en usage. | oui | oui | Troubles de la reproduction, immunosuppression, effets sur les glandes surrénales et thyroïdiennes. |
| BPC Biphényles polychlorés |
Transformateur électrique et fluide de condensateur, usage restreint en Amérique du Nord, mais pénètre dans l’environnement par les eaux de ruissellement, les déversements accidentels et l’incinération. | oui | oui | Troubles de la reproduction, anomalies squelettiques, immunotoxicité et perturbations endocriniennes. |
| Dioxines et furanes | Sous-produit du blanchiment au chlore, de la fabrication du bois et de la combustion incomplète. Les usines représentent maintenant une source moins importante. Les sources courantes comprennent la combustion de bois chargé de sel, les incinérateurs municipaux, la combustion résidentielle de bois de chauffage et de déchets de bois, les eaux de ruissellement provenant des boues d’épuration et le traitement du bois. | oui | oui | Dommages au thymus et au foie, anomalies congénitales, troubles de la reproduction, perturbations endocriniennes, immunotoxicité et cancer. |
| HAP Hydrocarbures aromatiques polycycliques persistants |
Sous-produit de la combustion de carburant, fusion de l’aluminium, traitement du bois, déversements accidentels d’hydrocarbures, usines métallurgiques et cokeries, usines de pâtes et papiers. | oui | non | Cancérogène. |
| Ignifuges, surtout PBB et PBDE Éthers diphényliques polybromés |
Ignifuges; dans les composants électriques et boîtiers de téléviseurs et d’ordinateurs, dans le textile et les rembourrages dans les automobiles, très répandu dans l’environnement. Deux des trois PBDE sont bannis en Europe. Les mêmes deux produits ont été retirés du marché en Amérique du Nord en 2005, mais un produit (décaPBDE) est encore utilisé à l’échelle mondiale. | oui | oui | Perturbations endocriniennes, dommages au foie et à la thyroïde. |
| PFO Perfluro-octane sulfonate |
Agents antitaches, hydrofuges et oléofuges (présent dans le Scotchgard jusqu’à tout récemment), mousse extinctrice, ignifuges, insecticides et frigorigènes, très répandu dans l’environnement. | oui | oui (dans le sang, le foie, les reins et les muscles) | Favorise la croissance de tumeurs. |
| TBT, DBT Tributylétain Dibutylétain |
Pesticide (peinture antisalissures) utilisé sur les navires. | oui | oui | Inconnu, mais associé récemment à une perte de l’ouïe. |
| PCP Paraffines polychlorées |
Ignifuges, plastifiants, peintures, agents de scellement et additifs utilisés dans les huiles lubrifiantes. | oui | oui | Perturbations endocriniennes. |
Organismes qui participent actuellement à des recherches sur l’otarie de Steller, population de l’est du Pacifique.
- Pêches et Océans Canada, Nanaimo, C.-B.
- Secteur des Sciences
- Marine Mammal Response Network
- Agence Parcs Canada
- Vancouver Aquarium Marine Sciences Centre, Vancouver, C.-B.
- Veterinary Department
- Marine Mammal Rescue and Rehabilitation Department
- Université de la Colombie-Britannique
- U.S. National Oceanographic and Atmospheric Administration, Seattle, Washington
- National Marine Fisheries Service
- Oregon Department of Fish and Wildlife
- Washington Department of Fish and Wildlife
- Alaska Department of Fish and Game
- Oregon State University
- Ministère de l’Agriculture et des Terres de la C.-B., Abbotsford, C.-B.
- Chercheurs indépendants
L’otarie de Steller du l’est du Pacifique est inscrite en tant qu’« espèce préoccupante » à l’annexe I de la Loi sur les espèces en péril (LEP). En tant qu’espèce aquatique, l’otarie de Steller relève de la compétence du gouvernement fédéral et est gérée par Pêches et Océans Canada (MPO), 200 -– 401, rue Burrard, Vancouver, Colombie-Britannique, V6C 3S4.
Le MPO a constitué un petit groupe de travail interne composé d’experts techniques provenant du domaine des sciences et de la gestion afin d’élaborer une première ébauche du présent plan de gestion. Un atelier technique sur la planification de la gestion de l’otarie de Steller a eu lieu en novembre 2007 et a servi de tribune où l’on a partagé des connaissances et de l’expertise sur les otaries de Steller, population de l’est du Pacifique, afin de soutenir l’élaboration du plan. Un groupe d’experts scientifiques et techniques, comprenant notamment des chercheurs indépendants, des organismes non gouvernementaux voués à l’environnement et d’autres membres des gouvernements fédéraux et provinciaux du Canada et des États-Unis ont participé à l’atelier. On a également envoyé une lettre d’invitation à l’ensemble des Premières nations côtières pour solliciter leur participation à l’atelier. Cet atelier a été très utile au groupe de travail interne du MPO pour la rédaction du Plan de gestion de l’otarie de Steller au Canada. Comme la population visée par le présent plan de gestion fréquente les eaux canadiennes et américaines, on a tenté d’obtenir une participation et une collaboration bilatérales gouvernementales et non gouvernementales.
Une version provisoire du plan de gestion a été publiée sur le site Web du MPO, Région du Pacifique, pendant une période de 30 jours (du 7 avril au 12 mai 2008) au cours de laquelle le public a pu la commenter; en outre, on a fait des envois postaux ainsi que des suivis par courriel à toutes les Premières nations vivant sur la côte afin de solliciter leur avis. Les consultations ont été faites en ligne et comprenaient une version initiale du plan de gestion, un guide de discussion et un formulaire de rétroaction en ligne. En outre, un message annonçant l’élaboration du plan de gestion a été envoyé à un serveur de liste (MARMAM) spécialisé dans les mammifères marins et assurant une vaste diffusion locale et internationale auprès de spécialistes des mammifères marins ainsi que de parties intéressées ainsi qu’à une liste de diffusion de personnes ressources en lien avec les baleines qui avait été fournie au MPO ces dernières années par des groupes environnementaux, des organisations non gouvernementales, des agences gouvernementales et le secteur de l’écotourisme.
Des commentaires relatifs au plan de gestion ont été reçus de huit sources indépendantes et de deux agences gouvernementales, à savoir Environnement Canada et le gouvernement de la C.-B. Les processus utilisés pour la coordination et la consultation entre les gouvernements fédéral et provinciaux concernant la gestion et la protection des espèces en péril sont exposés dans l’Accord Canada-C.-B. sur les espèces en péril (2005). Ressources naturelles Canada, le ministère de la Défense nationale, l’Agence Parcs Canada et Transports Canada n’ont pas commenté le document provisoire. L’Agence Parcs Canada a contribué à l’élaboration du plan de gestion en participant aux ateliers techniques et dans le cadre de ses fonctions de représentant, en tant qu’agence compétente en vertu de la LEP. Il n’était donc pas utile qu’elle prenne part au mécanisme de consultation publique. Par ailleurs, aucune rétroaction n’a été reçue à la suite des envois postaux destinés aux Premières nations.
Les commentaires formulés par le public, les agences gouvernementales et les scientifiques ont été examinés avec soin et pris en considération dans la production de la version finale du plan de gestion. Aucun examen par des pairs du document n’a été considéré nécessaire du fait que des experts en la matière ont participé à l’atelier technique sur la planification de la gestion de l’otarie de Steller et ont eu l’occasion de formuler leurs commentaires dans le cadre de la consultation publique.
Équipe technique de Pêches et Océans Canada
Paul Cottrell Pêches et Océans Canada
Marilyn Joyce Pêches et Océans Canada
Peter Olesiuk Pêches et Océans Canada
John Ford Pêches et Océans Canada
Graeme Ellis Pêches et Océans Canada
Jake Schweigert Pêches et Océans Canada
Peter Ross Pêches et Océans Canada
Larry Paike Pêches et Océans Canada
Tatiana Lee Pêches et Océans Canada
Joy Hillier Pêches et Océans Canada
Jeff Grout Pêches et Océans Canada
Linda Nichol Pêches et Océans Canada
Robin Abernethy Pêches et Océans Canada
Participants à l’atelier technique sur la planification de la gestion de l’otarie de Steller
Alana Phillips Vancouver Aquarium Marine Science Centre, B.C. Cetacean Sightings Network
Andrew Trites Université de la Colombie-Britannique
Annely Greene Pêches et Océans Canada, Gestion des pêches
Brent Norberg National Oceanic and Atmospheric Administration
Brian Fadely National Oceanic and Atmospheric Administration
Courtney Druce Pêches et Océans Canada, Gestion des pêches
Dave Rosen Université de la Colombie-Britannique, Marine Mammal Research Unit
Dominic Tollit Université de la Colombie-Britannique
Doug Bifford Gouvernement de la Colombie-Britannique
Graeme Ellis Pêches et Océans Canada – Région du Pacifique, Sciences
Jake Schweigert Pêches et Océans Canada – Région du Pacifique, Sciences
John Ford Pêches et Océans Canada – Région du Pacifique, Sciences
Joy Hillier Pêches et Océans Canada – Région du Pacifique, Habitat
Kaja Brix National Oceanic and Atmospheric Administration, Juneau, Alaska
Kim Raum-Suryan Alaska Department of Fish and Game et Université d’État de l’Oregon
Larry Paike Pêches et Océans Canada, Conservation et protection
Louvi Nurse Pêches et Océans Canada, Direction générale des traités et des politiques autochtones
Marilyn Joyce Pêches et Océans Canada, Gestion des pêches
Marty Haulena Vancouver Aquarium Marine Sciences Centre
Norm Sloan Agence Parcs Canada, Haïda Gwaii
Pat Gearin National Oceanic and Atmospheric Administration
Peter Ross Pêches et Océans Canada – Région du Pacifique, Sciences
Robin Abernethy Pêches et Océans Canada – Région du Pacifique, Sciences
Robin Brown Oregon Department of Fish and Wildlife
Tatiana Lee Pêches et Océans Canada – Région du Pacifique
Wendy Szaniszlo Agence Parcs Canada
1 Les contaminants préoccupants réglementés et non réglementés sont énumérés à l’annexe II.
2 La grande majorité des cas d’emprisonnement de pinnipèdes (110 sur 170) peuvent être associés à des dispositifs de protection contre les requins utilisés à un site; ces dispositifs ont été enlevés depuis (MPO, données non publiées).
3 Par exemple : Loi sur la marine marchande du Canada, Règlement sur la prévention de la pollution par les navires et sur les produits chimiques dangereux.
4 Inclure dans le manuel opérationnel les mesures précisées dans le guide de Pêches et Océans Canada sur les interventions en cas d’incident en lien avec les mammifères marins et le document de travail intitulé « Sea otter oil spill response plan for Canada’s Pacific coast » (plan d’intervention en cas de déversement d’hydrocarbures pour la loutre de mer de la côte canadienne du Pacifique).
5 Les agences gouvernementales et organisations non gouvernementales sont présentées à titre indicatif, et leur désignation n’oblige pas celles-ci à mettre en œuvre la mesure inscrite. La mise en œuvre des mesures sera fonction des priorités et des contraintes budgétaires de chaque organisme ou agence.
Détails de la page
- Date de modification :